Our recent focus on habitability addresses a significant problem. In order for astrobiologists to home in on the best targets for current and future telescopes, we need to be able to prioritize them in terms of the likelihood for life. I’ve often commented on how lazily the word ‘habitable’ is used in the popular press, but it’s likewise striking that its usage varies widely in the scientific literature. Alex Tolley today looks at a new paper offering a quantitative way to assess these matters, but the issues are thorny indeed. We lack, for instance, an accepted definition of life itself, and when discussing what can emerge on distant worlds, we sometimes choose different sets of variables. How closely do our assumptions track our own terrestrial model, and when may this not be applicable? Alex goes through the possibilities and offers some of his own as the hunt for an acceptable methodology continues.
by Alex Tolley
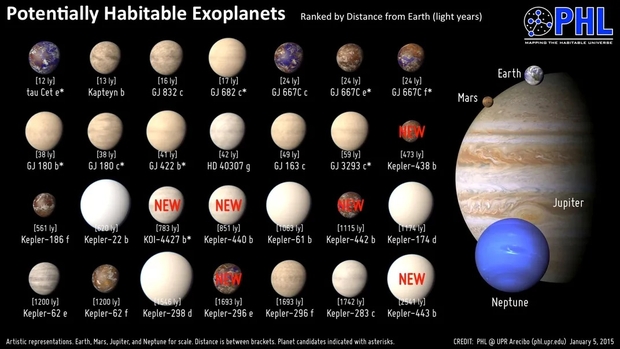
Artist illustrations of explanets in the habitable zone as of 2015. None appear to be illustrated as possible hanbitable worlds. This has changed in the last decade. Credit: PHL @ UPR Arecibo (phl.upr.edu) January 5, 2015 Source [1]
We now know that the galaxy is full of exoplanets, and many systems have rocky planets in their habitable zones (HZ). So how should we prioritize our searches to maximize our resources to confirm extraterrestrial life?
A new paper by Dániel Apai and colleagues of the Quantitative Habitability Science Working Group, a group within the Nexus for Exoplanet System Science (NExSS) initiative, looks at the problems hindering our quest to prioritize searches of the many possible life-bearing worlds discovered to date and continuing to be discovered with new telescope instruments.
The authors state that the problem we face in the search for life is:
“A critical step is the identification and characterization of potential habitats, both to guide the search and to interpret its results. However, a well-accepted, self-consistent, flexible, and quantitative terminology and method of assessment of habitability are lacking.”
The authors expend considerable space and effort itemizing the problems that have accrued: Astrobiologists and institutions have never defined “habitable” and “habitability” rigorously, even confounding “habitable” with “Habitable Zone” (HZ), and using “habitable” and “inhabited” almost interchangeably. They argue that this creates problems for astrobiologists when trying to plan how to develop strategies when determining which exoplanets are worth investigating and how. Therefore, defining terminology is important to avoid confusion. As the authors point out, researchers often assume that planets in the HZ should be habitable and those outside its boundaries uninhabitable, even though both assertions are untrue.
[I am not clear that the first assertion is claimed without caveats, for example, the planet must be rocky and not a gaseous world, such as a mini-Neptune.]
As our knowledge of exoplanets is data poor, it may not be possible to define whether a planet is habitable based on the available information, which leads to the imprecision of the term “habitable”. In addition, not only has “habitable” not been well-defined, but neither have the requirements for life been defined, which is more restrictive than the loose requirement for surface liquid water.
Ultimately, the root of the problem that hampers the community’s efforts to converge on a definition for habitability is that habitability depends on the requirements for life, and we do not have a widely agreed-upon definition for life.
The authors accept that a universal definition of life may not be possible, but that we can, however, determine the habitat requirements of particular forms of life.
The authors’ preferred solution is to model habitability with joint probability assessments of planetary conditions with already acquired data, and extended with new data. This retains some flexibility in the use of the term “habitable” in the light of new data.
Figure 1 below illustrates the various qualitative approaches to defining habitability. Adhering to any single definition is not possible for a universal definition. The paper suggests that a better approach is to use quantitative methods that are both rigorous, yet flexible in the light of new data and information.
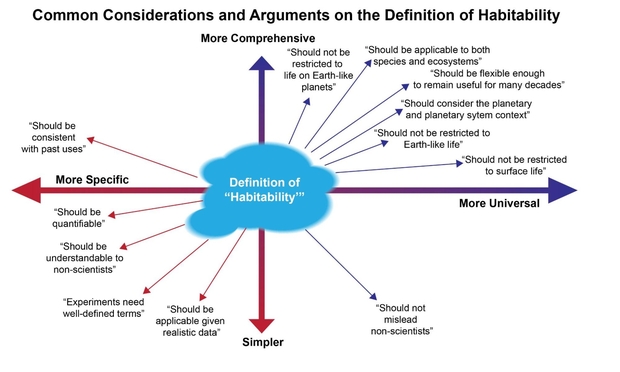
Figure 1. Various approaches to defining “habitable”. Any single definition for “habitability” fails to meet the majority of the requirements.
The equation below illustrates the idea of quantifying the probability of a planet’s habitability as a joint probability of the known criteria:
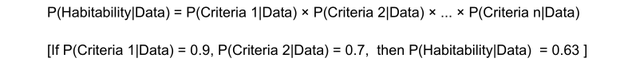
The authors use the joint probability of the planet being habitable. For example, it is in the HZ, is rocky, and has water vapor in the atmosphere. Clearly, under this approach, if water vapour cannot be detected, the probability of habitability declines to zero.
Perhaps of even greater importance, the group also looks at habitability based on whether a planet supports the requirements for known examples of terrestrial life, whose requirements vary considerably. For example, is there sufficient energy to support life? If there is no useful light from the parent star in the habitat, energy must be supplied by geological processes, leading to the likelihood that only anaerobic chemotrophs could live under those conditions, for example, as hypothesized in dark, glacial-covered subsurface oceans..
The authors include more carefully defined terms, including: Earth-like life. Rocky Planet, Earth-Sized Planet, Earth-Like Planet, Habitable Zone, Metabolisms, Viability Model, Suitable Habitat for X, and lastly Habitat Suitability which they defined as:
The measure of the overlap between the necessary environmental conditions for a metabolism and environmental conditions in the habitat.
Because of the probability that life (at least some species of terrestrial life) will inhabit a planet, the authors suggest a framework where the Venn diagram of the probability of a planet being habitable intersects with the probability that the requirements are met for specific species of terrestrial life.
The Quantitative Habitability Framework (QHF) is shown in Figure 2 below.
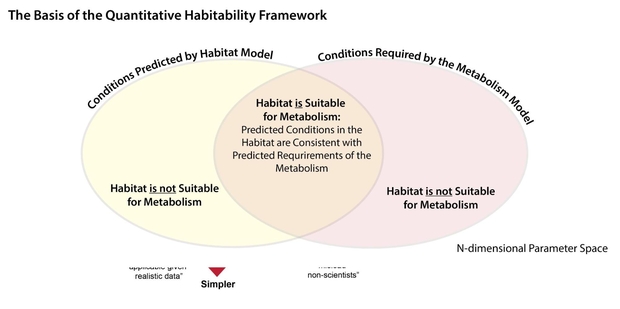
Figure 2. Illustration of the basis of the Framework for Habitability: The comparison of the environmental conditions predicted by the habitat model and the environmental conditions required by the metabolism model.
The probability of the viability of an organism for each variable is a binary value of 0 for non-viability and 1 for viability. For archaea and temperature, this is:
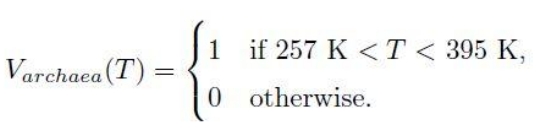
The equation means that the probability of viability of the archaea at temperature T in degrees Kelvin is 1 if the temperature is between 257 K ( -16 °C) and 395 K (122 °C). Otherwise 0 if the temperature is outside the viable range. (Ironically, this is imprecise, as it is for a species of archaean methanogen extremophile, not all archaeans.)
This approach is applied to other variables. If any variable probability is zero, the joint probability of viability becomes 0.
The figure below shows various terrestrial organism types, mostly unicellular, with their known temperature ranges for survival. The model therefore allows for some terrestrial organisms to be extant on an exoplanet, whilst others would not survive.
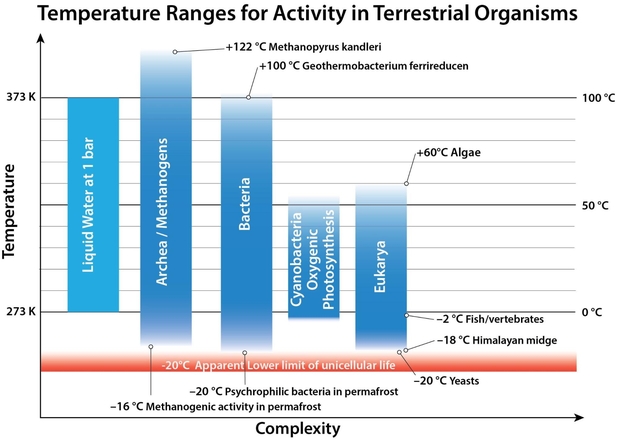
Figure 3. Examples of temperature ranges for different types of organisms, taken for species at the extreme ranges of survival in the laboratory.
To demonstrate their framework, they work through models for archaean life on Trappist 1e and Trappist 1f, cyanobacteria on the same 2 planets, methanogens (that would include archaea) in the subsurface of Mars, and the subsurface ocean of Enceladus.
Figure 4 below shows the simplified model for archaea on Trappist 1e. The values and standard deviations used for the priors are not all explained in the text. For example, the mean surface pressure is set at 5 Bar for illustrative purposes, as no atmosphere has been detected for Trappist-1e. The network model for the various modules that determine the viability of an archaean is mapped in a). Only 2 variables, surface temperature and pressure, determine viability of the archaean prokaryotes in the modeled surface temperature. In more sophisticated models, this would be a multidimensional plot perhaps using principal component analysis (PCA) to show a 3D plot. The various assumed (prior) and calculated (result) values and their assumed distributions are shown as charts in b). The plot of viability (1) and non-viability (0) for surface temperature and pressure is shown in the 3D plot c. The distributions indicate the probability of suitability of the habitat.
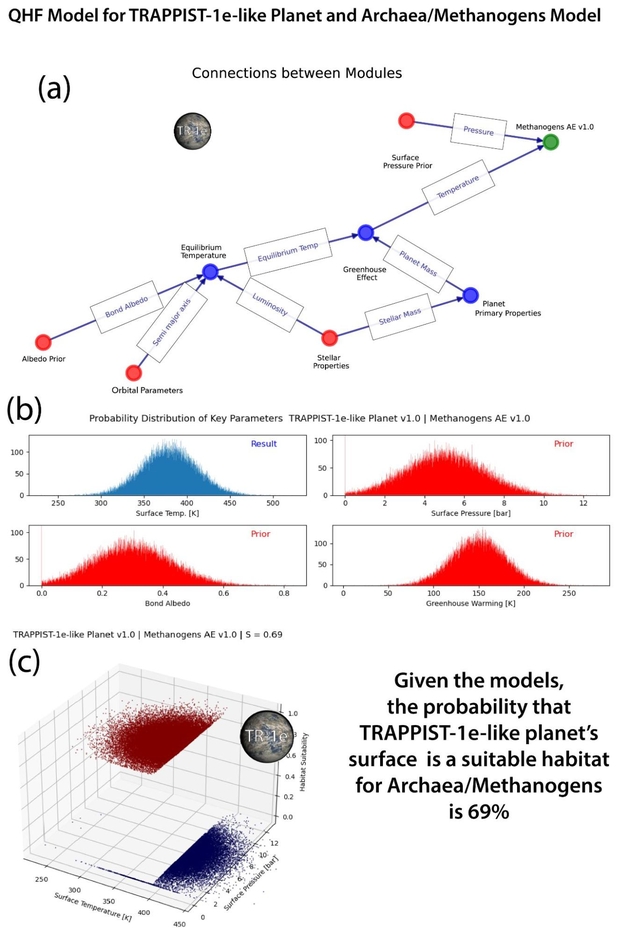
Figure 4. QHF assessment of the viability of archaea/methanogens in a modeled TRAPPIST-1e-like planet’s surface habitat. a: Connections between the model modules. Red are priors, blue are calculated values, green is the viability model. b: Relative probability distributions of key parameters. c: The distribution of calculated viability as a function of surface temperature and pressure. The sharp temperature cutoff at 395K separates the habitat as viable or not.
The examples are then tabulated to show the probabilities of different unicellular life inhabiting the various example worlds.
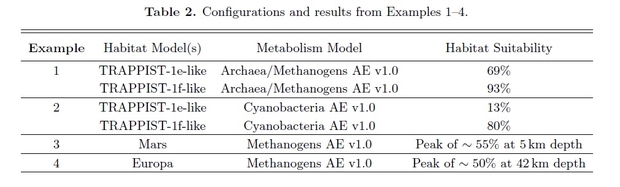
As you can see, the archaea/methanogens on Trappist 1f have the highest probability of being present inhabiting that world if we assume terrestrial life represents good examples. Therefore, Trappist 1f would be prioritized given the information currently available. If spectral data suggested that there was no water in Trappist 1f’s atmosphere, this would reduce, and possibly eliminate, this world’s habitability probability, and with it the probability of it meeting the requirement of archaean life and hence reducing the overlap in the habitability and life requirement terms to nil.
My Critique of the Methodology
The value of this paper is that it goes beyond the usual “Is the exoplanet habitable?” with the usual caveats about habitability that apply under certain conditions, usually atmospheric pressure and composition. The habitable zone (HZ) around a star is calculated for the range of distances from the star where, with an ideal atmosphere composition and density, on a rocky surface, liquid water could be found. Thus, early Mars, with a denser atmosphere, could be habitable [2], and indeed, the evidence is that water was once present on the surface. Venus might also once have been habitable, positioned at the inner edge of our sun’s HZ, before a runaway greenhouse made the planet uninhabitable.
The concept of NASA’s “Follow the water” mantra is a first step, but this paper then points out this is only part of the equation when deciding the priority of expending resources on observing a prospective exoplanet for life. The Earth once had an anoxic atmosphere, making Lovelock’s early idea that gases in disequilibrium would indicate life, which quickly became interpreted as free oxygen (O2) and methane (CH4), largely irrelevant during this period before oxygenic photosynthesis changed the composition of the atmosphere.
Yet Earth was living within a few hundred million years after its birth, with organisms that predated the archaea and bacteria kingdoms [4]. Archaea are often methanogens, releasing CH4 into the early atmosphere at a rate exceeding that of geological serpentinization. Their habitat in the oceans must have been sufficiently temperate, albeit some are thermophilic, living in water up to 122 centigrade but under pressure to prevent boiling. If the habitability calculations include the important variables, then their methodology offers a rigorous way to determine the probability of particular terrestrial life on a prospective exoplanet.
The problem is whether the important variables are included. As we see with Venus, if the atmosphere was still Earthlike, then it might well be a prospective target. Therefore, an exoplanet on the inner edge of its star’s HZ might need to have its atmosphere modeled for stability, given the age of its star, to determine whether the atmosphere could still be earthlike and therefore support liquid water on the surface.
However, there may be other variables that we have repeatedly discussed on this website. Is the star stable or does it flare frequently? Does the star emit hard UV and X-rays that would destroy life on the surface by destroying organic molecules? Is the star’s spectrum suited for supporting photosynthesis, and if not, does it allow or prevent chemotrophs to survive? For complex life, is a large moon needed to keep the rotational axis relatively stable to prevent climate zones and circulation patterns from changing too drastically? Is the planet tidally locked, and if so, can life exist at the terminator, as we have no terrestrial examples to evaluate? The Ramirez paper [3] includes his modeling of Trappist 1e, using the expected synchronization of its rotation and orbital period, resulting in permanently hot and cold hemispheres.
While the authors suggest that the analysis can extend beyond species to ecosystems, and perhaps a biosphere, we really don’t know what the relevant variables are in most cases. Unicellular organisms are sometimes easily cultured in a laboratory, but most are not. We just don’t know what conditions they need, and whether these conditions exist on the exoplanet. It may be that the equation variables may be quite large, making the analyses too unwieldy to be worth doing to evaluate the probability of some terrestrial life form inhabiting the exoplanet.
A further critique is that organisms rarely can exist as pure cultures except in a laboratory setting with ideal culture media. Organisms in the wild exist in ecosystems, where different organisms contribute to the survival of others. For example, bacterial biofilms often comprise different species in layers allowing for different habitats to be supported, from anaerobes to aerobes.
The analysis gamely looks at life below the surface, such as lithophilic life 5 km below the surface of Mars, or ocean life in the subsurface Enceladan ocean. But even if the probability in either case was 100% that life was present, both environments are inaccessible compared to other determinants of life that we can observe with our telescopes. This would apply to icy moons of giant exoplanets, even if future landers established that life existed in both Europan and Enceladan subsurface oceans.
What about exoplanets that are not in near circular orbits, but more eccentric, like Brian Aldiss’ fictional “Heliconia”? How to evaluate their habitability? Or circumbinary planets where the 2 stars are creating differing instellation patterns as the planet orbits its close binary?. Lastly, can tidally locked exoplanets support life only at the terminator that supports the range of their known requirements, such as Ramirez’ modeling of Trappist 1e?
An average surface temperature does not cover either the extremes, for example the tropics and the poles, nor that water is at its densest at 277K (4 °C), ensuring that there is liquid water even when the surface is fully glaciated above an ocean. Interior heat can also ensure liquid water below an icy surface, and tidal heating can contribute to heating even on moons that have exhausted their radioactive elements. If the Gaia hypothesis is correct, then life can alter a planet to support life even under adverse conditions, stabilizing the biosphere environment. The range of surface temperatures is covered by the Gaussian distribution of temperatures as shown in Figure 4 b.
Lastly, while the joint probability model with Monte Carlo simulation to estimate the probability of an organism or ecosystem inhabiting the exoplanet is a relatively computationally lightweight model, it may not be the right approach with more variables added to the mix. The probabilities may be disjoint with a union of different subsets of variables with joint probabilities. In other words, rather than “and” intersections of planet and organism requirement probabilities, there may be an “or” union of probabilities. The modeled approach may prove brittle and fail, a known problem of such models, which can be alleviated to some extent by using only subsets of the variables. Another problem I foresee is that a planet with richer observational data may score more poorly than a planet with few data-supported variables, simply due to the joint probability model.
All of which makes me wonder if the approach really solves the terminology issue to prioritize exoplanet life searches, especially if a planet is both habitable and potentially inhabited. It is highly terrestrial-centric, as we would expect, as we have no other life to evaluate. If we find another life on Earth, as posited by Paul Davies’ “Shadow Biosphere,” [4] this methodology could be extended. But we cannot even determine the requirements of extinct animals and plants that have no living relatives, but flourished in earlier periods on Earth. Which species survived when Earth was a hothouse in the Carboniferous, or below the ice during the global glaciations? Where were those conditions outside the range of extant species? For example, post-glacial humans could not survive during the Eocene thermal maximum.
For me, this all boils down to whether this method can usefully help determine whether an exoplanet is worth observing for life. If an initial observation ruled out an atmosphere like any of those Earth has experienced in the last 4.5 billion years, should the search for life be immediately redirected to the next best target, or should further data be collected, perhaps to look for gases in disequilibrium? While I wouldn’t bet that Seager’s ‘MorningStar’ mission to look for life on Venus will find anything, if it did turn up microbes in the acidic atmosphere’s temperate zone, that would add a whole new set of possible organism requirements to evaluate, making Venus-like exoplanets viable targets for life searches. If we eventually find life on exoplanets with widely varying conditions, with ranges outside of terrestrial life, would the habitat analyses then have to test all known life from a catalog of planetary conditions?
But suppose this strategy fails, and we cannot detect life, for various reasons, including instrumentation limits? Then we should fall back on the method I last posted on, which reduces the probability of extant life on an exoplanet, but leaves open the possibility that life will eventually be detected.
The paper is Apai et al (2025)., “A Terminology and Quantitative Framework for Assessing the Habitability of Solar System and Extraterrestrial Worlds,” in press at Planetary Science Journal. Abstract.
References
1. Schulze-Makuch, D. (2015) Astronomers Just Doubled the Number of Potentially Habitable Planets. Smithsonian Magazine, 14 January 2015.
https://www.smithsonianmag.com/air-space-magazine/astronomers-just-doubled-number-potentially-habitable-planets-180953898/
2. Seager, S. (2013) “Exoplanet Habitability,” Science 340, 577.
doi: 10.1126/science.1232226
3. Ramirez R., (2024). “A New 2D Energy Balance Model for Simulating the Climates of Rapidly and Slowly Rotating Terrestrial Planets,” The Planetary Science Journal 5:2 (17pp), January 2024
https://doi.org/10.3847/PSJ/ad0729
4. Tolley, A. (2024) “Our Earliest Ancestor Appeared Soon After Earth Formed” https://www.centauri-dreams.org/2024/08/28/our-earliest-ancestor-appeared-soon-after-earth-formed
5. Davies P. C. W. (2011) “Searching for a shadow biosphere on Earth as a test of the ‘cosmic imperative,” Phil. Trans. R. Soc. A.369624–632 http://doi.org/10.1098/rsta.2010.0235



This discussion is very interesting and offers ways to make progress in making strategies for directing our search for extra terrestrial life. I think it would be a great help to setting search strategies if someone could demonstrate experimentally at least one path from inorganic chemistry and environs to organic chemistry and then to cell-based life. Given the habitat ranges of extremeophiles on Earth, perhaps there is more than one pathway starting point (not a single one that adapted over time) and it would be good to demonstrate that too.
I agree with Alex’s critique and I can identify items to add to the list! Maybe more if I were to read the paper.
Ultimately I doubt that we can unambiguously declare an exoplanet habitable until we discover that it is in fact inhabited. And vice versa. We’re swimming in a sea of ignorance (what is habitable, life, etc.) that continues to be almost entirely sterile due to the lack of data. That includes ignorance about which exoplanets are fruitful to study, if study them we can. I suspect that this paper doesn’t really help.
@Ron.
Conceptually, they have a sensible idea in trying to quantify the probability of habitability, by being more precise with terminology and especially which terrestrial life forms could be extant. That should, theoretically, improve resource use. In practice, I agree with your sentiment. We are far too ignorant about life. Their analysis assumes that life will appear quickly so that there is no “Great Filter” early on in a planet’s history. As we only have knowledge of terrestrial life, they have to use that as the canonical life forms for their analysis. Until we have data from a good sample of worlds about habitability and inhabited worlds, this analysis method may prove useless as a quantifier for the probability of life on any given exoplanet. What I do think might happen is that astrobiologists will use this method to justify their chosen planet for observation, even if the method really doesn’t have much to say about life, especially in hindsight.
It is rather like the Drake Equation that lends a sense of quantification of the probability of receiving signals from a civilization. In practice, our knowledge of the evolution of life, intelligence, and technological, communicating civilizations is so poor that one can posit that we are alone in the galaxy, to a galaxy positively brimming with such civilizations eager to communicate. The number of civilizations the group came up with justified using radio telescope resources to look for signals. I suspect motivated reasoning, even though I think it was, and is, worth pursuing.
Regarding SETL, what I would prefer is more, and hopefully better, approaches to remotely detect life, whether in our system and especially on exoplanets.
As a reader and follower of yours and other constructors of early life scenarios, it takes a while to appreciate or comprehend many of the well reasoned elements.
But looking at the same problem from a different perspective, and since it still causes some disarray, that habitability or habitable zone controversy, there is a criterion or measure that I think could help.
My Johnny One Note idea for exoplanet life’s prospects:
Use the stellar effective temperature at orbital radius as a measure for habitability, not the calculated exoplanet temperature subject to … Well, everything.
Compared to exoplanets, stellar surface temperatures are very stable. The less massive the star, the longer it remains so. For the sun right now and at the standoff distance of the Earth’s radius, effective surface temperature is equivalent to 395 K, or less than 400 K, vs. the 5800 K or so of the solar surface. My surmise is that this relation has not changed much over geological periods of time.
But on the other hand, the Earth’s surface temperature has. It has iced over, its atmospheric chemical constituents have changed and so has its albedo or reflectance. Partly due to the changing chemical constituents, prominence of clouds at various layers, ocean and ice caps, and chemical emissions of life itself.
Even the forms of life have changed atmospheric and ocean albedo.
To take a more extreme case, Venus argues against many HZ’s that would nominally include it. Maybe it could have better qualified during some yesterday.
While one star’s mass and surface temperature results in a different overall blackbody surface temperature, a radius for 350 or 400 K radial effective temperatures still can be derived. The less massive the star, the longer this
relation will prevail as observed, but at closer radius.
Then, of course, the red dwarfs, as we have often discussed here, have a large number of emissions that make that 400 K radius rather dangerous. Maybe the K and F regions of the main sequence should be examined more closely on that account.
And I would suspect that some extrapolations would be easier than others from our own history, 9/10ths of which appears to have been without
much diatomic oxygen in our atmosphere. So much of what is discussed above is extrapolation from what can be discerned about the earliest era of life here, a tunnel out of which we so recently emerged by comparison.
I should note that despite not being stars or even brown dwarfs, gas giants and “Neptunes” in our solar system have atmospheric profiles that will allow 300 or 400 kelvin temperatures in something like an ocean of atmosphere. Somewhere in their depths. Had Titan been orbiting closer to the sun we would be even more excited about its rather dense atmosphere. How devoid of atmosphere it would be by comparison if it were closer to the sun does not necessarily preclude exoplanets that could look like Triton in a more habitable state.
But as a life abode, why NOT consider gas giants? Probably because extracting information from beneath their clouds is difficult enough in this solar system, never mind beyond. But I would bet that they are chemical cauldrons.
Strangely though, despite Galilean and other systems, we have not discovered as yet any corresponding exoplanet systems of moons. But if a gas giant resides at a 300 to 400 K effective temperature radius from an MS star of sufficient lifetime, why shouldn’t there be a moon and a prospect for life – somewhere in the system?
Perhaps the tightly bound HZs of the easily observable exoplanet transits about red dwarfs make it difficult to establish 3 body systems involving moons. But elsewhere that should not be as problematic. However, gathering transit data on Earth with our moon for LGM would take years, literally.
For terrestrial planets vs. gas giants, it would appear that the preceding biochemistry would perhaps have some roots in the stellar nursery gas cloud.
The chemical complexity of these regions seems to be increasing. The mechanisms for transporting precursors to the surfaces of planets like Earth appears both arduous, but evidently often successful, judging from some
interpretation of asteroid sample retrievals. Gas giants, however, it seems like
they would each represent a abiogenesis case of their own, owing to the difficulty of any precursor biochemistry surviving a comet impact. Yet could a jovian atmosphere ( somewhere?) be akin to the million monkeys at typewriters pecking toward a Shakespeare play? If life were to get out and we would inquire of it where it came from. The answer: “You would never guess…”
So the 400K value must have been lower when the sun was 30% fainter than it is today. It will be higher in about 1 billion years. But if we believe life existed on Earth within a few hundred million years of its formation, and should last another billion years on land, and longer in the oceans, is that value or range of values that useful?
We could debate that.
Life (as we know it) would not have appeared as a separate abiogenesis on Jupiter. Maybe life could live there now due to panspermia in the past. Even weirder if Earth is providing a constant flux of microbes to Jupiter to maintain a population in its atmosphere. As you know, the Vesian atmosphere has a temperate zone tens of km from the surface. Dray, highly acidic, but Seager still thinks it is worth looking for life there. I think the anomalies detected will be due to chemistry, but I endorse her looking.
I think we have explored the issue of how wet a planet can be at formation, from very dry (like the fictional Arrakis – “Dune”) all the way to hycean worlds. If we “follow the water,” then all wet worlds are habitable by that criterion.
While Earth has suffered both global ice ages, and extreme heat (the end of the Permian), both geology and life act as temperature stabilizers, even if human life is currently destabilizing the temperature as did the cyanobacteria, but we are doing it over centuries rather than tens or hundreds of millions of years.
As we don’t know about any life beyond Earth, we have no idea if terrestrial life is THE way for life to exist, or one of very many alternatives that may occupy conditions that would be impossible for terrestrial life. As a result, we are forced to look for our life. Because we cannot conceive of FTL starflight that is closer to terrestrial travel times both past and present, there is no hope that we can study life forms on exoplanets with scientific exploration ships. What we might be able to do eventually is build vast, high-resolution telescopes that can provide sufficient imaging to truly detect life directly on some exoplanets, without being trapped by false positives like the “vegetation on Mars”. That will be a long way off, but such [swarm] telescopes are physically feasible, and perhaps more useful than small telescopes at the SGL. We will see what technology and economics can manage.
I believe there are some very good ideas for life detection by proxy, but they require far higher resolution spectroscopy to achieve. Whether they can also detect “life as we don’t know it”, assuming it could exist, I don’t have a good grasp of that.
A.T.,
Your point about zero main sequence age luminosity is well taken.
The 400 K region, in the early days, would be moved away from Earth – and toward Venus.
A little less than half a magnitude. Where I have seen this feature invoked is in terrestrial studies with our own sun. Since main sequence formation behavior is not uniform, I am not sure how this rule of thumb would work out with a K, an M or an F. And getting more familiar with this particular article, it looks like Trappist-1 ( M9 or so) system is the main illustrative example.
Paradoxically as well, it was billions of years ago that Mars is suspected of having had large surface bodies of water and the atmospheric circulation that
would entail.
Yet even as a star in the main sequence moves slightly ( horizontally and or vertically) on the main sequence, my point is that the stars behavior is more stable than the orbiting planets. In early epochs, planets have heat of formation leaking out much more profusely than they will a billion years or more later. And if they are like Earth in having water, as the planet cools, the subsurface boiling point will change as well as the atmospheric observed state.
Reflected light away from the atmosphere and absorption into it vary considerably for complex reasons without life being part of the atmospheric chemistry. But in terrestrial geological history, sometimes it is due to the
anaerobic life forms. Distinguishing between chemical and biological atmospheric transformations, I would infer, is underway even now.
But if we are assuming surface conditions conducive to life around 300 Kelvin or overall slightly above freezing, a planet’s geothermal history could be more turbulent than the radiative flux through space that stars overhead provide.
Midwestern US temperatures in a year might vary between 255 to 310 K on a given day. That’s due to the seasons, day-night and weather; not so much the solar variation or even orbit eccentricity. Yet “climate” factors such as locally induced climate change can make local or planetary temperatures a further departure from this without the sun doing anything different.
What might be an interesting distinction in this argument: suspected exoplanet climate variations due to the presence of primitive life or proliferation of precursors. On Earth we could assume that there was once a time when oceans were devoid of life. Hence, they would provide certain features if observed from space.
… Ruminations on your proposition, of course. But it does seem like there ought to be some threshold exoplanets out there that are nearly the blank slates that life would become established on. If the boundary between no life and life could be more sharply determined for observation programs, maybe
we can spot exoplanets just about ready to start and those which have just left the starting blocks. Trappist-1 planets are rather elderly. But if they never got a living start, then their more detailed characterization might give some clues
of the “before”, assuming they never reached the “after”.
WDK
I think the Ramirez paper in the references includes the model that allows for Mars to be warm and wet after its formation, allowing surface oceans/seas. You might want to read that section.
Both heat from radio isotopes (e.g., Al_26), plus the heat released from bombardment, keep a planet warm while the star is fainter. You may recall a CD article I wrote about internal heat allowing liquid water below the surface for subsurface biospheres to exist on worlds where the surface is below freezing.
I would like to see some way of detecting directly whether a planet is inhabited or not. I am skeptical it can be done because we cannot even do this from the surface and close orbit of Mars. For Earth-sized planets with an atmosphere, that may be a different matter. Here we are back to atmospheric gas disequilibria, trace biosignature gases, and with high-resolution images, the detection of large areas of plants on land, and even in the oceans.
There are ways to rule out false positives and that is where the Earth sized planet in the Goldilocks zone pays off if there is oxygen in the atmosphere. A G class star helps. We know that photosynthesis produces oxygen. The problem is getting the spectra over one hundred light years away which is the limit of JWST. The ESO ELT is completed in 2028 does not fare any better. What if the nearest exoplanet with oxygen is over 400 light years away? New techniques to improve old technology will be helpful.
@Geoffrey
We really cannot be certain that free O2 in the atmosphere is not due to photolysis. That is why we still need to detect other gases in disequilibrium with the O2 to be more confident it is due to oxygenic photosynthesis. What if life exists, but does not have such photosynthesis, or just reduces sulphur compounds like green bacteria?
I would like to see other proxies that can “triangulate” the life case by ruling out abiotic processes, possibly very exotic ones.
The more we look for an “Earth 2.0” world, the more likely it seems that we will detect life, assuming that it is not vanishingly rare. OTOH, if we find many terrestrial worlds that look like Earth in its early Archaean stage, then if we ever manage to travel to those worlds using some technique to carry whatever we call humanity in that period, then we could colonize them without fear of disrupting their non-existent biospheres. [By then, spacefaring humanity may not even be biological, the biologicals remaining on Earth.]
As you say, we need better instruments. However, I will say that we have made remarkable progress in my lifetime. Just watch old sci-fi movies to realize how far we have come, and the continuing rapid progress we are making as we bring more computational power to bear.
We have not seen any false positives in Earth sized exoplanets in the life belt around any stars. We could use the other criteria to eliminate the false positives, the water vapor and biosignature gases like carbon dioxide, methane, etc. to rule the false positives out. We’ve done that already with hot Jupiters, etc. where we know that they are most likely false positives.
I completely agree with the Earth 2.0 idea as there is a great deal of contingencies, but the question is do all of them apply to the Earth 2.0 in order for it to have life. Intelligent life certainly must be contingent on the Earth 2.0 model and moving away from them the less chances of intelligent life. There still must be contingencies for microbial life to evolve.
An Earth 2.0 model does make life rare, but there still could be hundreds of Earth 2.0’s in our galaxy. The ethics of colonization don’t apply as long as our presence does not interfere with the planets natural evolution. If it does it could be considered unethical. I have to agree with that approach as opposed to one which throws the baby out with the bathwater. In other words, I think the Earth 2.0 model eventually will be proven to be correct based on the first principles of physics and physical science.
In the spirit of the quantifying model, how would you define “Earth 2.0”?
The authors define an Earth-like Planet:
“Earth-2.0” would be more or less defined than Earth-like in your opinion? If needing more definition, what would you add, and perhaps what variables with ranges of values would you want to add to make the definition “Earth 2.0”? If more loosely defined than “Earth-like”, what variables would you expect in “Earth-like” to be dropped?
The size of the planet which is not enough. We must use our Earth as the 2.0 model and include all contingencies that sustain life over deep time. So far our planet is the most habitable. We include type of solar system that Earth is in a G class star. It must have gas giants in the outer solar system at least one large on like Jupiter with a strong gravity to deflect meteorites. It is crucial for the planet to have a Moon based on the giant impact hypothesis, so the Earth 2.0 can have a fast rotation to have a magnetic field. The Moon also is involved with tidal forces and plate movement in the crust in plate tectonics. The planet has to be in the life belt. Having an Earth like solar system might be a necessity due to general relativity and gravitational effects keeping the system in balance. This is speculative of course, but my point here is to illustrate that there are hidden contingencies that are not considered that all add up to be important and yes make life more rare at least intelligent life, so the Earth 2.0 idea has to be thought of as part of a larger system.
The idea of the Earth 2.0 model is a way to rule out the oxygen false positive. For example. If the most important contingencies of the Earth 2.0 model are met, then we know that the probability of abiotic oxygen in the atmosphere is much lower than biotic oxygen. It does not mean there is not any life without oxygen in the spectra, but we have to go there to prove it. This is why it is so important to start with the Earth 2.0 model which is all we have and supported completely by first principles.
ET’s who are hundreds of thousands of years more advanced than us in our galaxy have the capability to see the spectra of oxygen in our atmosphere with huge space telescopes and know that the potential for life being very high here, the potential for intelligent life depending on the age of our Star. The oxygen in our atmosphere has been in the visible spectrum for at least half a billion years and longer.
So, apart from the large moon (hard to detect even now), and the need to do the search for O2/O3, are there any candidates in the current exoplanet catalog that meet your criteria to target with a spectrographic search for confirmation of a favorable atmosphere for aerobic life?
While I do think this model gives us a tool to help us understand the problem, it is ultimately not helpful. The term habitability needs to be scrapped at least when it comes to life on planets.
The terms comes from the title of Stephen Doles’ book “Habitable Planets for Man” in which he analyzes planets for human settlement, a mindset derived from our colonizing settlement patterns on Earth. The field of exoplanetology has since grow to such an extent that analyzing planets for their habitability is like analyzing the field of medicine using the terminology of the four humors. It’s redundant. And aside from which Dole was specially aiming for man-habitability.
What we need to be doing is developing models for planet types that can give rise to life, planet types that can give rise to complex, macroscopic life, and they are stable for long enough that sentient life may arise and we get a technological civilization. (These planets may all be completely uninhabitable for man—if they have 5 bars of Nitrogen in their atmosphere.)
If we are looking for habitable planets for man, life man not even be involved. The is a chance (a faint chance) that photolysis will provide the right level of Oxygen for the atmosphere to be breathable despite the planet being sterile as the general chemistry is too oxidizing for life to arise.
There is one other point I’d like to make. To quote: “For example, it is in the HZ, is rocky, and has water vapor in the atmosphere. Clearly, under this approach, if water vapour cannot be detected, the probability of habitability declines to zero.”
This is almost certainly wrong. The spectra come from the 1 mb level (175,000 ft on Earth) and at that height water vapor can be completely frozen out. The JWST got spectra from K2-18b and TOI-412b. K2-18b and TOI-412b are similar planets both in size and chemistry, but K2-18b showed no water lines while, TOI-412b, which is warmer, did.
@Dave
I think that it is a strawman argument to use Dole’s use of the word from a book explicitly talking about worlds to settle, while astrobiologists use it in a different context.
I almost disagree with every part of this statement.
1. abiogenesis. Yes, we should assume that is the case, but I wouldn’t rule out panspermia, especially for worlds in the same system
2. Complex life. On Earth, complex life didn’t appear for at least 3 bny, and possibly nearly 4 bny. This suggests that most planets like Earth will only be inhabited by simple life.
3. Sentience and technology. While SETI assumes this is likely, I would be very careful in making that assumption. Furthermore, humanity might conceivably wipe itself out, which would imply that any SETI attempt might be futile if intelligence and technology prove to be self-eradicating.
IDK enough about the technology, but I would be surprised that transmission spectroscopy of the atmosphere would fail to detect water vapor somewhere in the atmosphere column. If we could detect the spectrum reflected off the planet rather than when it was in transit, wouldn’t the absorption from the surface water be evident? [As would the “red edge” of terrestrial chlorophyll in the oceans and on the land.]
This opening sentence is the one I would tend to agree with…with caveats (weasel word, I know).
On the plus side of your comment, it reminded me to have another look for Dole’s book. The RAND ppbk (2007) can be freely downloaded as a PDF file, but I also found a near fine copy (1970) with dust jacket for a reasonable price, and ordered it for my home library. [I am a sucker for early pop science books that have become dated. Fred Hoyle’s astronomy books are marvelously written, but sometimes end with incorrect deductions – most famously the “Steady State” theory.]
FWIW I saw an online interview with a retired professor who when in graduate school had encountered some astronomers who still accepted the steady state theory. He said they kept their jobs and came into their offices every day, they just didn’t have any students. So it may be that science advances not one funeral but one retirement at a time…felt a bit sorry for them though…
@NS
While I believe the Steady State model is dead, the studentless tenured professors may have the last laugh as some evidence suggests the Guth Big Bang model may not be correct either. There seem to be suggestions that the universe was created from a prior universe.
I do wonder what happens to professors who stake their reputations on a theory that gets falsified. Do they stay on drawing a salary, but get no students for postgraduate work? Do they focus of different areas of their discipline? Or do they take early retirement and leave academia, perhaps attending dinners and meetings to keep up relationships?
I’ll vote for “planetary and planetary system context”. Neptune’s ocean is much too hot for life – perhaps if it is ejected from the system it might cool down enough? Mercury’s poles are at a near-ideal temperature, if you can get around the issue of an atmosphere. Saturn, Venus, and Jupiter all have habitable zones in the atmosphere, depending on your pH preferences. Ceres bears the hallmarks of a warm ocean once upon a time. Earth offers liquid water and ideal conditions for life, provided you don’t dawdle more than a century on the way over: we’re almost to an end of the century level of heating now, and the moist greenhouse can’t be too far off. It seems somehow satisfying to imagine that you can’t really predict if a planet can host life, until after you’ve explored it.
That may be correct, but more in a moment. But consider. You are an astrobiologist. ET life has not yet been found. There is fame for you in the history books if you are the first. There are several possible worlds in our system, and thousands of possible exoplanets to search. How do you become the first?
The best proof would be a sample from our system, culled from Mars or an icy moon. It would be pretty definitive, and likely satisfy the 2nd genesis or panspermia question. But this is a long shot. Our system may be sterile beyond Earth, planetary landers are expensive long-term projects, and the present US administration is intent on gutting NASA and especially space science projects, even those currently operating. So probably better to go for an analysis of the telescope data, especially telescopes that cannot be stopped from operating by US funding cuts. So, which target do you want to pick? Your approach needs to compete for access and time with other astrobiologists and astronomers for time on these telescopes. Probably the best chance to find life is to determine which is the most likely Earth-like planet within the set, which is close enough to get good quality spectral data. Do you try for JWST time, with NIR & IR detection, or perhaps a terrestrial telescope with better spectrometry, or do you work with a team to build a better, higher-resolution spectrometer, perhaps with an intent to launch a small private telescope with that device?
My sense is that the quantification method to assess probabilities will be part of your analysis, both to hopefully increase your chances of being first, but also to get your proposal funded.
Now, on to the issue of life itself. Life may be ubiquitous and appear on any suitable planet. That would be the best hope for detection, although, as on Earth, many suitable worlds may be in the aquatic prokaryotic organism stage. (Maybe you look for older stars that have had a longer time for evolution.) Perhaps life is very rare. The probability of an inhabited planet, however apparently suited, is so low that it is just serendipity if you pick an inhabited world. All that Bayesian analysis of null detection shows that if/when life is unambiguously positively detected, it is an exceedingly rare event, and the statistics don’t tell you much about the requirements for life that result in its occurrence. It just appears to be almost random.
In this latter case, even if you could mount crewed starflight missions in FTL ships and surveyed perhaps a thousand worlds, if perhaps 5 or 10 proved inhabited, yet the other 990-995 proved sterile yet had the same conditions as the inhabited world, would that provide you with more information on where to find life, or not? My sense is that your intuition may be correct, and why I quoted your last sentence.
At this point, we don’t know anything about ET life, and are just crossing our fingers that we will be successful once our instruments are good enough to detect it, even if just by proxy. If the only ET life is beyond our system, then we won’t have any wriggling, motile organisms under a microscope, nor even the more imaginative images of macroscopic life as depicted in Wayne Barlowe’s “Expedition: Being an Account in Words and Artwork of the 2358 A.D. Voyage to Darwin IV”. I just hope I live long enough to see the first highly probable detection of exoplanet life.
@Alex: I understand that this perspective makes sense for those close to the action. But from an outsider’s point of view, I’m seeing the Planetary Society say there is a 47% cut in funding. Even projects with an obvious and pragmatic rationale, such as prospecting the Moon and Mars, are to be lost. It’s hard to picture that risky SETI research should be able to compete under those circumstances, even with a ‘better’ chance of finding life. There is a more detailed breakdown at AAS, with some interesting points noted in an interview at WSWS (now outdated), as the cuts were twice as large).
Though there is good reason to think of this as a single catastrophic event, I think we should view this in a broader context. American presidents have been promising to go to Mars “in a decade” every decade since the Moon landings. Privatization appeared to put the Moon within reach, but it amounted to a contest between two companies: one expensive and not innovative, the other ambitious … and run by a CEO who took only five years to go from his first launch carrying astronauts to the ISS to threatening to strand them there (barring rescue by friendly Russians) if the government didn’t give him free money for other ventures. Meanwhile, the slow and steady Moon Rabbit seems bound to win the race: Chang’e 7 should launch next year, and by 2028 a Chinese mission will be addressing the practical problems of doing robotic manufacturing and construction on the Moon.
Thanks for the AAS link on budgets. Who made those budget decisions, and why? The sums are so small as to be almost meaningless, yet the science cost is high. Was it to stick it to those “pointy-headed” science elites? Or just a chainsaw to make it easier to justify the tax cuts by introducing distractions?
AFAICS, the Chinese robots are not yet as sophisticated as the NASA robots on Mars. But I approve of their use, and we can see the many failures of other institutions to successfully put a lander on the Moon, including Russia which did successfully do over 50 years ago. Lunokhod program.
Aren’t the Chinese indicating that they want to put astronauts on the Moon in the 2030s?
Some AI help has turned up interesting information. Space is mentioned briefly in the 14th Five Year Plan (2021-2025):
“We will perform basic scientific research on the origin and evolution of the universe and perspectives on the earth (透视地球), carry out interstellar exploration such as Mars orbiting and asteroid inspection, develop a new generation of heavy-lift launch vehicles and reusable space transportation systems, deep earth exploration equipment, deep sea operations and maintenance (O&M) and equipment test ships, polar three-dimensional monitoring platforms, and heavy icebreakers, and complete construction of Phase IV of the lunar exploration project, Phase II of Jiaolong Sea Exploration (蛟龙探海), and Phase II of Xuelong Polar Exploration (雪龙探极).”
The AI thinks that Phase IV extends to the initial phases of construction of the International Lunar Research Station; I didn’t see explicit sources for that, but all indications are that the existing program is meant to set the stage for it to happen. So far, descriptions of the Chinese moon program seem strangely similar to those of the old American program, but given the careful efforts being made to develop robotic technology to build the base, and the accompanying explorations of harsh environments on Earth, it seems like the program would be handled differently in practice. I imagine intriguing detail will be released in association with the publication of the fifteenth Plan by next year. The strategic importance of controlling the rim of Shackleton Crater, beginning with the Chang’e 7 mission next year, clear: the U.S. had planned to land on the same territory.
The restriction of parameter space to look for Life As We Know It makes sense, in the same way that the proverbial drunk looking under the lamp-post for his keys makes sense. But it’s an admission of failure of imagination to look no further afield.
Disequilibrium gases seem a more agnostic approach based on first principles, but they’re prone to a bit too much bias from observers invoking abiotic processes as a default rather than as an option on equal footing.
@Adam
Going back to Lovelock’s gases disequilibrium is still the easient to do with spectroscopy, even if it has been added to with so-called biosignature gases.
The problem I have with disequilibrium is with the classic methane-oxygen disequilibrium. We know that the serpentinization reaction also creates CH4. Because of this, any CH4/O2 disequilibrium relies on the amount of CH4 generated by methanogens to greatly exceeed that of geologic processes. Can we rely on terrestrial data to apply to all exoplanets, or does it mean that the disequilibrium detected must be considered a proxy biosignature with some probability attached?
[Why can’t the boffins devise an FTL propulsion system like in sci-fi? LOL]
Reading through articles linked to this theme of ET life, the analogy of the drunk only looking for their keys under the lamp post is very apt. Indeed it is apt to pretty much all walks of like, from cancer research to rocket design to devising “good” political strategies. It is in many ways how science works – we build on the data we have or can generate.
We have no data beyond Earth, i.e. one data point, one lamp post. Barring something like Arrival or Independence Day or a visit from a Volcan, all distal observations will ultimately fall short of broad crediblility.
Our easiest path to a clear answer is to go to Mars and spend a couple of decades there. It is the best chance we have to adding to our singular data point.
Already have – with robotic flybys, orbiters, and landers. The issue is whether humans on Mars is worth the cost, and whether they can do something that rovers cannot – like drill deep cores, retrieve samples, and analyze the samples for life. Are we better off waiting for more capable robots to do the work, or trying to send humans to Mars for a real scientific expedition sometime in the 2030s – 2040s? I would like to see robots set up a drill site and manage to drill down to 1 km, and retrieve a sample, which is analyzed for various organic compounds. This isn’t easy, as was made plain by a recent human deep drilling in the Antarctic to reach a subsurface lake. Robots are not yet capable of that task. However, given how fast robots are improving, maybe that is the better approach given the resources. It certainly is cheaper than a human expedition, and given the US’ current science cuts, possibly more likely to be achieved by another nation.
Already have – with robotic flybys, orbiters, and landers. The issue is whether humans on Mars is worth the cost, and whether they can do something that rovers cannot – like drill deep cores, retrieve samples, and analyze the samples for life.
I meant for people, lots of people, to be there and a log or three increase in the spend. With robots, it is like fusion, it’ll be here in “20-years’ time”.
Would it be worth it? This is tricky. For me it is the most pressing question of our time – are we alone? Establishing a second genesis on Mars or remains of a similar genesis would point to a fertile universe. Nothing there would point to a barren one. A few or even an army of robots would in many ways be less efficient than letting people get to work there for a decade or two.
NASA’s current annual budget is ~ $25bn.
US current GDP ~ $28tn
Obviously, this 3 order magnitude spending to put loads of people on Mars is a non-starter for the USA.
World GDP ~ $110tn
Spending 1/4 of global GDP on putting lots of people on Mars is also a non-starter. Even if it were somehow possible, the cost really isn’t worth the gain. We cannot even send $100bn to poor countries to help mitigate climate change. I would rather spend the billions, if not trillions of $s, countering climate change, saving ecosystems and biodiversity as much as possible, both on land and in the oceans. Possibly finding evidence of Martian life, extant or extinct (and preferably a 2nd abiogenesis) is not worth doing if we impoverish the one planet we know has a rich biosphere undergoing a 6th major extinction event.
It might take 20-50 years for robots to be able to erect a rig to do deep drilling on Mars, and repeat the exercise a number of times around Mars, but I would rather do this since the spinoff technology to reach that stage of robot competency to work in teams like humans, would be immense. Robots, whether humanoid or not, are far better adapted to working on Mars for years, needing only access to electrical energy to maintain themselves (apart from accidental damage). The rovers have demonstrated this already.
We can send robots and a lot of equipment to Mars as slowly as we want with any chosen propulsion system[s]. They are literally “hibernating” on the journey and are woken when they land on Mars. They needn’t return. So much easier than sending humans who will need LSS to reach Mars, and while on Mars, and the ability to return. And we do this because today only humans can erect drilling rigs (but can they in spacesuits?).
I think it’s safe to say that the Mars part of that budget is significantly south of 25 billion. By “Mars part”, I would mean that specific to getting there, staying there and so forth.
We pretty much have the tech to go there and stay there. We need the desire. Pussy footing around will get is there “in 20”.
Regarding the robots, I’m reminded of the recent clip of them playing football…