Centauri Dreams
Imagining and Planning Interstellar Exploration
Thinking about Bracewell Probes
Sometimes I jog my perspective on thorny physics issues by going back to earlier work. At our all too infrequent dinners together, Claudio Maccone used to tease me about this, saying that older scientific papers had inevitably been superseded by recent work which would, in any case, incorporate the early documents. But I find that looking at an idea afresh sometimes means re-living its inception, which puts things in context. It was in that spirit that I recently revisited a key paper by Ronald Bracewell.

The name Bracewell holds a certain magic, invoking as it does the era when SETI was just beginning and speculations about extraterrestrial civilizations were getting wider circulation outside the science fiction magazines. Bracewell (1921-2007) was Australian by birth, acquiring degrees in mathematics and engineering and joining in work on World War II era radar. Following completion of a PhD in physics at Cambridge, he continued his work in the 1950s with a position as senior research officer at the Radiophysics Laboratory of the Commonwealth Scientific and Industrial Research Organisation (CSIRO).
Image: Ronald N. Bracewell, Stanford, CA, March 1983. Credit: NRAO/AUI Archives, Sullivan Collection. Located through Wikimedia Commons.
Bracewell came to the U.S. in 1954 to lecture on radio astronomy at UC-Berkeley before joining the Electrical Engineering department at Stanford University. His contributions to interferometry and the calibration of radiotelescope instruments to achieve breakthrough results are substantial, as a quick look through NASA’s Astrophysics Data System under his name reveals. I’ve noticed in scanning through this body of work that his interest in interstellar probes was persistent as he continued to contribute to the science of exoplanet discovery.
Nestled within the ADS results from 1960 is the unusual paper titled “Communications from Superior Galactic Communities,” which ran in Nature in 1960. In this early era, we had just had the famous paper from Giuseppe Cocconi and Philip Morrison (citation below) that is widely regarded as the beginning of modern attempts to find extraterrestrial civilizations. Given that this paper ran in Nature, which Bracewell obviously knew well because he was writing for it, we can assume that Cocconi and Morrison triggered his decision to write about the SETI question.
I call SETI a ‘question’ in this case because what struck Bracewell about it was its impracticability. Remember, at this same time, Frank Drake had begun planning (in 1959) for the project that would become Ozma, listening to Tau Ceti and Epsilon Eridani in 1960, and it’s evident that Cocconi and Morrison spurred the conference at Green Bank in 1961 that led to the creation of the famous Drake Equation. So we are witnessing western (as opposed to Soviet, with its somewhat different perspectives) SETI beginning to emerge, and it seemed to Bracewell that its approach was off-center.
This comes across in the “Communications from Superior Galactic Communities” paper loud and clear. Going through the suggestion from Cocconi and Morrison that 1420 MHz was the ‘waterhole’ frequency around which radio-using civilizations in search of an audience would gather, Bracewell then mentions Drake’s plans, and points out how unlikely it is that ETI would find us. After all, we were at that time looking for a radio beacon singling us out:
Let us assume that there are one thousand likely stars within the same range as the nearest superior community. This makes it hard for us to select the right one. Furthermore, if this advanced society is looking for us, we can only expect to find them expending such effort as they could afford to expend on the thousand likely stars within the same range of them. It does not seem likely that they would maintain a thousand transmitters at powers well above the megawatt estimated by Drake as a minimum for spanning only 10 light years, and run them for many years, and we could scarcely count on them paying special attention to us. Remember that throughout most of tho thousands of millions of years of the Earth’s existence such attention would have been fruitless.
The alternative? Send probes to nearby stars designed to attract the attention of technological beings on any planets there. It is indicative of the optimism of the early space era that Bracewell should describe interstellar flight as “…what we ourselves are now discussing and are on the point of doing, probably during this century…” We now look to the possibility of an interstellar probe by the end of this century, but the physics says the idea is doable.
Unlike SETI, where we cope with the inverse-square problem of attenuation of the signal, we would be talking about a probe within no more than a few minutes or hours of communications time from its target. Travel times are obviously lengthy, but with an eye toward science delivery for coming generations Bracewell suggests ‘swarm’ strategies that would deliver probes to perhaps the thousand stars near enough to us to be of interest. Each probe could quickly learn key facts about life and technology on these worlds.

Image: Ronald Bracewell (left), with Stanford’s Von Eshleman, a major figure in early research into gravitational lensing. Here the two are examining the horn antennae that Bracewell used in 1969 to determine that the Sun is moving relative to the cosmic background radiation. Credit: Linda Cicero/Stanford University.
In 1974, Bracewell would investigate the prospect of a galactic ‘network’ of civilizations, one that we could perhaps join, but even here in 1960 he homes in on the idea. He imagines our world joining a perhaps galaxy-spanning ‘chain of communication,’ and thus dealing with civilizations that have been through the contact scenario many times on many worlds. These would, obviously, be superior technologies from which we could learn new science.
Bracewell’s probes, then, are designed for contact, and meant to be identified by ETI. He would expand these ideas in his 1974 book The Galactic Club: Intelligent Life in Outer Space. The version of this title most likely to be available in used book stores is the 1976 printing from the San Francisco Book Company, and it’s a good thing for any interstellar enthusiast to track down.
Here the method reminds us that not long after the time Bracewell was writing, Carl Sagan was negotiating with Russian astronomer Iosif S. Shklovskii to reprint the book that would become in its western edition Intelligent Life in the Universe (Holden-Day, 1966). The story of that collaboration is itself interesting, as Shklovskii didn’t realize Sagan would not just publish his book Universe, Life, Mind in the west, but would also heavily annotate it with his own brand of science popularization. That disharmony apart, Sagan’s awareness of Bracewell becomes apparent given the method of communications that ETI uses with Earth to announce their presence in the novel Contact, the re-broadcast of radio messages from our past.
Bracewell had suggested something similar, though using radio:
Such a probe may be here now, in our solar system, trying to make its presence known to us. For this purpose a radio transmitter would seem essential. On what wave-length would it transmit, and how should we decode its signal ? To ensure use of a wave-length that could both penetrate our ionosphere and be in a band certain to be in use, the probe could first listen for our signals and then repeat them back. To us, its signals would have the appearance of echoes having delays of seconds or minutes such as were reported thirty years ago by Størmer and van der Pol and never explained.
I don’t want to get caught up in the famous delayed-echo story of the 1920s, but the short version is that amateur radio operator Jørgen Hals observed echoes of a Dutch shortwave station in 1927 and took the matter to Norwegian physicist Carl Størmer and Dutch physicist Balthasar van der Pol. The echoes became the subject of work by Scottish writer Duncan Lunan, who explored them as possible signs of a Bracewell probe operating in the Solar System. The claim became controversial, to say the least, and has since been refuted, although Lunan continued to investigate it. And it is also true that long-delayed echoes have been attributed to various natural sources but remain enigmatic.
In any case, Bracewell advocated remaining alert to a possible interstellar origin for signals that are unusual, for the benefits of joining in an interstellar conversation would be immense. He calculated that even if there were few civilizations that outlived their adolescence (remember, this was in the Cold War era, with nuclear destruction always on our minds), there might still be a few that survived and went on to long lifetimes. The paper continues:
Presumably such an ancient association would be very able indeed technically, and might seek us out by special means that we cannot guess. Whether they would be interested in rudimentary societies which, in their experience, would usually have burnt themselves out before they could be located and reached, is hard to say. Such communities would be collapsing at the rate of two a year (103 in 500 years), and they might already have satisfied the!r curiosity by archreological inspection made at leisure on sites nearer home. On the other hand, the prospect of catching a technology near its peak might be a strong incentive for them to reach us.
Bracewell’s place in the early SETI literature, including Michael Hart and Frank Tipler, can’t be examined without bringing in John von Neumann, whose self-reproducing machines would likewise have spurred Bracewell’s imagination, though his own concept did not include this capability. I want to try to fit some of these pieces together and likewise bring back Sagan and Shklovskii in the next essay. What we’re juggling here is the very concept of what Sagan called ‘mediocrity,’ which he described as ‘the idea that we are not unique.’ Do we sometimes stretch our Copernican understanding of the cosmos too far?
The paper is Bracewell, “Communications from Superior Galactic Communities,” Nature Volume 186, Issue 4726 (1960), pp. 670-671. Abstract. The Cocconi & Morrison paper is “Searching for Interstellar Communications,” Nature 184 (4690) (1959), pp. 844–846. Full text.

Interstellar Choices: Where to Look for Habitability?
A recent conversation with a friend who works the futures markets has me thinking about the nature of daydreaming. This is a guy who tracks fast-breaking numbers all day long so as to avoid getting a freight-car’s worth of coffee beans or some other commodity delivered to his condo. His numbers, he says, are all business, and allow no time for daydreaming. Whereas the numbers I study have no deadline, and give me plenty of time for reflection, moments of gazing off into the distance and just letting thoughts run. Today, for example, I’m troubled about what we know about the age of the galaxy.
If daydreaming sounds abstract, consider that this is an issue that has a bearing on our own standing in the cosmos. We have a pretty good read on the age of the Earth, and can peg it at around 4.5 billion years. Various sources tell me the Big Bang occurred some 13.8 billion years ago, with the formation of the Milky Way beginning not terribly long thereafter. Let’s say for the sake of argument that our galaxy is 13.6 billion years old, a figure that NASA recently cited.
So when did worlds like the Earth – terrestrial planets – began to appear? I think I’ve been writing about this question since Centauri Dreams first appeared, as it draws upon the work of Charles Lineweaver (Australian National University), who in 2001 landed on the figure of 9 billion years ago. The problem is immediately apparent: The galaxy seems to be stuffed with many a planet that is older than our own, and in many cases considerably so. Lineweaver’s work found that the median age of terrestrial planets is on the order of 6.4 billion years.
Here we tug again at the Fermi question – ‘Where are they?’ – since these numbers suggest that the opportunity for civilizations to emerge was robust long before our planet began to coalesce. Since that seminal 2001 paper, which I’m surprised is not cited more than it is, Lineweaver has continued to explore the numbers, and they are likewise massaged in other subsequent papers, but rather than going into the details, let’s just say that we’re still left with a galaxy far older than our planet. Give an extraterrestrial civilization a 2 billion year head start and you might think they would be visible to us in some way, or maybe not. Maybe civilizations don’t live all that long?
See Stephen Webb’s wonderfully readable If the Universe is Teeming with Aliens, Where is Everybody? (Springer 2015), the latest edition of which offers 75 answers to Fermi that range from the preposterous to the ingenious. I also send you to Milan Ćirković’s absorbing The Great Silence: Science and Philosophy of Fermi’s Paradox (Oxford, 2018), which mines the depths of a question that many do not consider a paradox, and others find deeply troubling no matter what the name. And Paul Davies is also a reminder of how rich the literature on Fermi is. See his The Eerie Silence (Mariner, 2010) for still further insights.
Thinking about a culture that was around in the days when the first signs of life began to appear on Earth is indeed cause for daydreaming. I notice this morning that Avi Loeb, in his lively publishing venture on Medium, is looking at how long-lived civilizations might cope with the problems raised by their longevity. It’s one thing to consider our own fate when the billion years or so we have before the Sun gets too hot to deal with completely dwarfs our species’ scant time on Earth. But what would we do if we actually survived for that billion years? Would we go elsewhere, or find a way to move the Earth to an orbit that would provide habitable conditions for millions, even billions of years more?
This is pretty lively stuff, for it opens up the possibility of terrestrial-class planets orbiting far outside what was once their habitable zone. It also brings into question the matter of white dwarfs, which could still sustain life for a species that insisted on staying within its natal stellar system. An ETI that can move planets might move one again, this time back in toward the Earth-sized remnant of its former red giant star. I would assume interstellar relocation would make more sense, but no one can know what alien minds might think of this.
Loeb has worked on these issues before:
In 2013, I co-authored a paper with Dani Maoz… which showed that during a transit by an Earth-mass planet across a white dwarf, the transmission spectrum of the planet’s atmosphere would show prominent bio-markers such as molecular oxygen absorption at a wavelength of ∼ 0.76 micrometers. We calculated that a potentially life-sustaining Earth-like planet transiting a white dwarf would be detectable by the Webb telescope in about 5 hours of total exposure time, integrated over 160 two-minute transits.
The method is familiar, one that we’ve discussed here often ever since the first transmission spectroscopy results began showing us what could be found in a hot Jupiter’s atmosphere. I love the idea of expanding the search for habitable worlds into environments as seemingly bizarre as these, although the limitations on telescope time (demand is high!) would make such searches lower priority than, say, a close look at a nearby red dwarf’s habitable zone planet. Here again we have more SF story material, though. All the possible planets around white and red dwarf stars make for fertile hunting for story crafters.

Image: Artist’s impression of a still unconfirmed planet around the white dwarf star WD1054-226 orbited by clouds of planetary debris. Credit Mark A. Garlick / markgarlick.com. License type Attribution (CC BY 4.0).
Loeb also mentions a paper I had missed in earlier discussions of stellar ages. In 2019, Nicholas Fantin (University of Victoria, BC) and colleagues extended the Lineweaver work I led this post with to include white dwarfs, considering them as age markers that help us trace the development of the galaxy. The bare bones of this method are described here:
We develop a new white dwarf population synthesis code that returns mock observations of the Galactic field white dwarf population for a given star formation history, while simultaneously taking into account the geometry of the Milky Way (MW), survey parameters, and selection effects. We use this model to derive the star formation histories of the thin disk, thick disk, and stellar halo.
Skipping the details, I just want to cite a few results that back up the interesting point about the relative youth of the Sun. According to this model, the Milky Way’s thick disk began forming stars 11.3 ± 0.5 billion years ago. The growth rate peaked at 9.8 ± 0.3 billion years ago. A slow decline in starbirth is traced that eventually became a constant rate that persists until now. Heavily reliant on results from the Gaia mission, the data set is dominated by disk stars in the solar neighborhood. A larger sample size will eventuate through surveys like Pan-STARRS DR2, the LSST, as well as data from WFIRST and Euclid.
Again we face what Tennyson called ‘the long result of time.’ So much time, in fact, that civilizations in their multitudes would have had the chance to form. Cirkovic notes in The Great Silence just how much deeper the Fermi question becomes when we consider it in light of such findings. He points out that the original Fermi statement (WeakFP) could be taken to ask why we have seen no evidence of extraterrestrials on Earth or in the Solar System. Keep extending the search outward, though, and the issue gets more and more puzzling. Take the entirety of our past light cone as your canvas and the lack of signs of extraterrestrial activity despite the billions of years civilizations could have existed escalates in impact. This is why Webb’s book is as long as it is.
All this is occurring even as we continue to rack up exoplanets of all descriptions including those of terrestrial mass, and even as the prospect of interstellar travel is now under serious investigation, as we’ve just been reminded by Jim Benford’s work with Breakthrough Starshot. We have developed, says Cirkovic:
Improved understanding of the feasibility of interstellar travel in the classical sense and in the more efficient form of sending inscribed matter packages over interstellar distances. The latter result is particularly important since it shows that contrary to the conventional skeptical wisdom shared by some of the SETI pioneers, it makes good sense to send (presumably extremely miniaturized) interstellar probes, even if only for the sake of communication.
Just where to send such probes? The nearest stars are obvious candidates, with Proxima Centauri b leading the list, but fleshing out a target roster – today an exercise in theory more than planning – may take in destinations we have only begun to consider. That’s assuming our early work on interstellar probe technologies continues to develop options for ever more distant targets. Imagine ‘swarm’ flybys of interesting systems, a capability we may well be able to deploy some time late in this century.
The nearest white dwarf to the Sun, by the way, is Sirius B, some 8.6 light-years out. The closest solitary white dwarf is van Maanen’s Star, about 14 light years distant. The closest red giant is Pollux in Gemini, at about 34 light years distance
The paper is Fantin et al., “The Canada-France Imaging Survey: Reconstructing the Milky Way Star Formation History from Its White Dwarf Population,” The Astrophysical Journal Vol. 887, No. 2 (17 December 2019), 148. Full text. Charles Lineweaver’s 2001 paper is “The Galactic Habitable Zone and the Age Distribution of Complex Life in the Milky Way,” Science Vol. 303, No. 5654 (2 January 2004), pp. 59-62, with abstract here.
Biology on a Rogue Planet’s Moon?
I don’t write science fiction, but I have several friends who think I do simply because I write about distant planets and futuristic ways to reach them. The boundary between SF and science has always fascinated me. I like to poke around in old magazines, most of them from the science fiction field, but a particular interest is magazines like Hugo Gernsback’s Science and Invention and Radio News, early 20th Century venues for fiction that dealt with science and preceded 1926’s Amazing Stories.
Astronomy and fiction have been mingling for a long time, but as we uncover startling exoplanets and posit theories that explain them, I’m usually wondering how quickly an SF writer will pick up on the latest work with a stunning new setting. Today’s paper offers another opportunity, as it presents the possibility that ‘rogue’ planets, wandering in the interstellar dark without a warming Sun, may support biology not on their surfaces but on any potential moons.

Image: Artist’s rendition of a Jupiter-sized rogue planet moving through interstellar space without any star. Scientists have been exploring the possibility of life on worlds warmed by internal heating alone. A new paper now looks at moons around such worlds and the processes that could keep them warm. Credit: JPL/Caltech.
Scientists involved with the German research network ORIGINS, working with researchers at the Max Planck Institute for Extraterrestrial Physics (near Munich) believe that large moons of free-floating planets can retain liquid water oceans for over 4 billion years because of the twin effects of dense hydrogen atmospheres and tidal heating. That closes in on the amount of time Earth has existed, with the obvious implication that complex life could develop.
Lead author David Dahlbüdding (Ludwig-Maximilians-Universitat Munchen) is lead author of the study:
“Our collaboration with the team of Professor Dieter Braun helped us recognize that the cradle of life does not necessarily require a sun. We discovered a clear connection between these distant moons and the early Earth, where high concentrations of hydrogen through asteroid impacts could have created the conditions for life.”
Recent work has shown that a gas giant ejected from its birth system could retain moons despite the gravitational encounter that would have forced it into interstellar space. Orbiting moons would be nudged into elliptical orbits by the event, but the resulting tidal forces between moon and planet are a blessing in disguise, in that they could generate enough frictional heat to maintain surface oceans. An atmosphere rich in hydrogen can also undergo ‘collision-induced absorption.’ in which thermal radiation is then retained by the atmosphere.
Earlier papers have examined rogue planet atmospheres heavy in CO2, where atmospheric collapse is a probability. But the researchers think hydrogen is far more interesting. From the paper:
The present-day Earth looks much different from the worlds presented here, which, with their thick hydrogen envelopes and possibly deep oceans, resemble a Hycean planet. Although usually in the sub-Neptune range, these worlds are prime candidates for the detection of life (Madhusudhan et al. 2021, 2023a,b). In their case, any tidal heating could conversely narrow the habitable zone (Livesey et al. 2025). Our small-scale Hycean worlds could provide relatively better conditions for life. Due to their (∼ 25%) lower gravity, high-pressure ices between a potential liquid water ocean and the rocky core would be less likely, allowing the ocean to receive essential nutrients (Cockell et al. 2024). Although, as Madhusudhan et al. (2023a) note, this represents only one possible source of these essential biological elements.
What to make of this? Extending the range of possible biology is always interesting, but the natural question is how we might actually observe such a system. Free-floating planets are a difficult enough catch without bringing potential moons into the mix. Gravitational microlensing offers a faint possibility, but here we’re dealing with chance encounters with background stars that are beyond our conceivable likelihood to predict. Although the authors mention transits of the host free-floating planet, this seems quite a reach. How do we know where to look, when their presence is unpredictable? The Roman Space Telescope should detect plenty of rogue planets, but the issue remains – a gravitationally microlensed event is by its nature unrepeatable.
I don’t want to downplay targeted searches for young rogue planets still throwing a good infrared signature in their adolescence. These we might actually detect through direct imaging if we scan nearby star clusters, so it’s not outside the realm of possibility to think we might get a rare transit of a moon. But the unlikeliness of such a detection means we may have to chalk this up as a fascinating theoretical result without observational consequences, at least at the present state of our technology.
Still, what an interesting landscape for a science fiction tale…
The paper is Dahlbüdding et al., “Habitability of Tidally Heated H2-Dominated Exomoons around Free-Floating Planets,” in process at Monthly Notices of the Royal Astronomical Society 24 February 2026 (full text).
Starshot Is a Success (Part II)
The second part of Jim Benford’s examination of Breakthrough Starshot concludes our look at the numerous issues advanced by Phase I of the project. Largely discounted in recent press coverage, the Starshot effort in fact completed a successful Phase I and left behind numerous papers that illuminate the path forward for interstellar flight. This is solid work on everything from laser arrays to metamaterials and the engineering of data return at light-year distances. Read on.
by James Benford
“I have learned to use the word ‘impossible’ with great caution.”
— Wernher Von Braun, after the lunar landing
In this second report, I will describe the major results of Starshot beginning with the mission scenario and then treating each major technical area in terms of how solutions have been resolved and issues retired. In Part 1, I described Phase 1 objectives.
One of the causes of Starshot results not being well-known publicly is that the Breakthrough Foundation has not publicized its events and results during most of its duration. After its completion, substantial reports have appeared, but are not commonly available to the public. There is a final report, but it has yet to be published. There are briefings by Harry Atwater at Breakthrough Discuss and the IRG in Montreal in 2023 [1,2].
The most detailed discussions are in the book Laser Propulsion in Space edited by Claude Phipps, with a system overview by Pete Worden and others, a description of his system model by Kevin Parkin and other aspects of directed energy in space by Philip Lubin, all in the one volume [3]. The Kevin Parkin article is particularly interesting because it contains fully worked-out examples of the possibility of future voyages of humans traveling to the stars in large >100 m sailcraft in future centuries. Note that there are many journal publications produced by Breakthrough Starshot. And there are many papers that have been published since Starshot was put on hold.
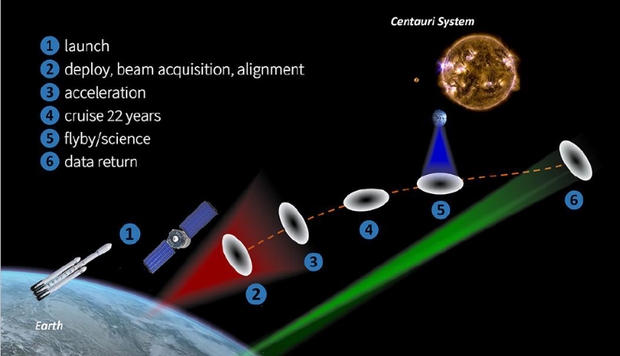
The Starshot Mission Scenario has evolved as a substantial improvement over previous beam-driven sail mission concepts. A mothership is launched which houses a fleet of membrane-like sailcraft measuring ~5 meters in diameter and less than a micron thick. The traditional standard laser guide star adaptive optic system can’t be scaled to Starshot-sized apertures to deal with the time–dependent fluctuations due to atmospheric turbulence. The system uses a satellite–based laser which is called the Beacon. It’s in an orbit at the launch time of apogee 200,000 km.
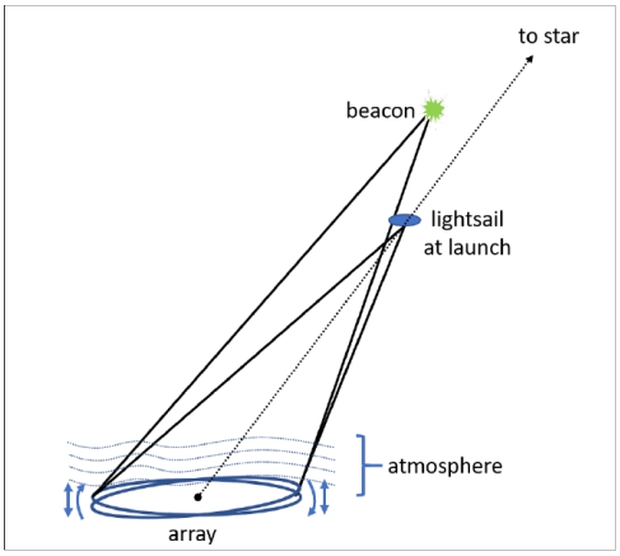
Image: Starshot system geometry. Arrows indicate that the array acquires atmospheric turbulence data from a Beacon and points the beam at the sailcraft. (Courtesy of Breakthrough Foundation.)
The sailcraft are composed of super-reflective metamaterials that stabilize the perturbations that could prevent beam-riding during the propulsion phase. The scientific instruments that are the payload are integrated into the sail. The mission begins as the mothership deploys a sailcraft into space.
Meanwhile on Earth, a phased array of 100 million small lasers turns on, generates ~100 GW of optical power and, using information from a Beacon in high orbit, digitally adjusts the phase of the emitted light to correct for atmospheric turbulence. These small lasers would be manufactured in printed sheets, following the fabrication techniques of the semiconductor industry. This is the means of lowering laser prices.
The single 100 GW beam focuses on the sail and accelerates the sail. Almost no energy is absorbed by the sail’s reflective surface, so imparting force. The sail rides the beam for ten minutes and reaches relativistic speed. It leaves the solar system in less than a week. Soon after acceleration it encounters dust and charged particles, so can be oriented edge-on to avoid such collisions. On arriving at the Alpha Centauri system, it captures images, detects dust and particles and measures fields. The sail transmits data home to an array of optical receivers on Earth, so it begins to arrive four years later. Data return may take decades because of limited data rate. Recall that complete data return from the New Horizons flyby took about a year.
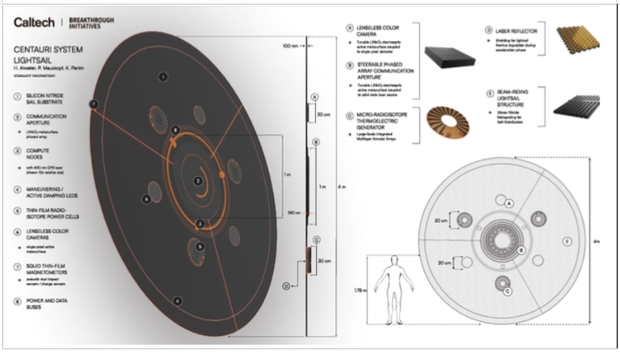
The above figure shows a concept for the sail, about 5 m in diameter. Some studies show that at the velocity under consideration the gas and dust will pass through the thin sail with virtually no damage if it travels face-on. Only the payload would need protection. The sail can also be oriented edge-on in order to avoid such collisions, giving meters of material protection to the center. The payload is around the center, protected from damage due to incoming gas and dust.
Key issues for beam-driven sail systems have been retired by high levels of Starshot research. Most are resolved at the conceptual level. Experiments are needed to verify solutions for these major issues, discussed below.
Can phase be maintained across a large aperture composed of many sources? This is well demonstrated historically for microwaves, principally for radar. For lasers, a new concept has been quantified [4, 5]. Building the hundred million laser emitters into a large array is the driving technical challenge of the project. The principle of the design is to interferometrically link multiple arrays which are phase-locked into modular tiers of larger size. That is, multiple areas which are individually phase controlled would be linked together by interferometry. This approach of linking multiple optical phased arrays is called a hierarchical array. The array design that resulted has laser dimensions and total power levels that are about five orders of magnitude beyond present state of the art capabilities. To control the phase over such a large aperture is the most significant technical challenge to Starshot.
Can a sail material be found which can meet the many constraints on sail acceleration? Most materials effort has been for laser propulsion, where the leading candidate for sail material is silicon nitride. There are no fundamental limits to optimize that material for the key parameters of mass, reflectivity, refractive index, and thermal properties. (For microwaves various types of carbon are preferred, such as microtruss and graphene.)
Can the sail ride on the beam stably? (Feedback is impossible over long ranges.) If not, sails can veer off-course on millisecond timescale. The notion of beam-riding, stable flight of a sail propelled by a beam, places considerable emphasis on the sail shape. Even for a steady beam, the sail can wander off if its shape becomes deformed or if it does not have enough spin to keep its angular momentum aligned with the beam direction in the face of perturbations. Beam pressure will keep a concave shape sail in tension, and it will resist sidewise motion if the beam moves off-center, as a sidewise restoring force restores it to its position. Early stability experiments verified that beam-riding does occur with a conical sail [6].

Experiments and simulations show that conical sails ride a microwave beam stably. The carbon–carbon sail diameter is 5 cm, height 2 cm, and mass 0.056 g.
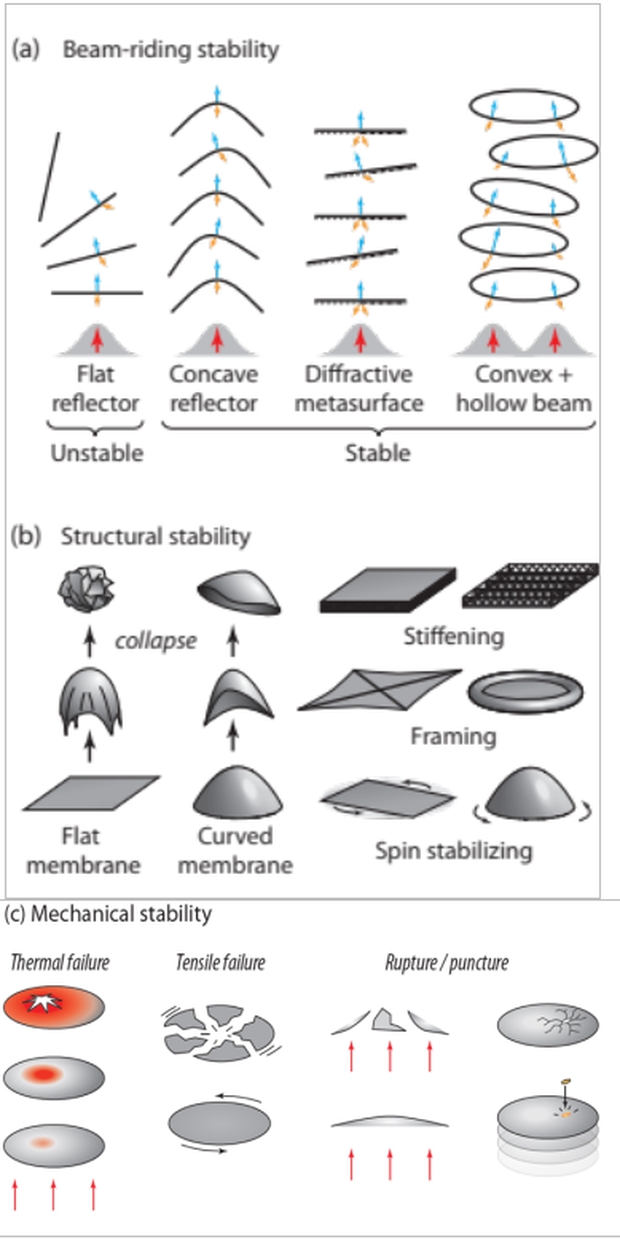
Beam riding and structural stability is difficult. (a), beam-riding stability, where bold upward arrows depict accelerating beam, light upward arrows the force of radiation pressure, downward arrows the direction of reflected light (b) structural stability methods (c) mechanical issues [7].
Meter-scale shaped sails of submicron, ~100 atomic layer thickness can ride with stability along the axis of the accelerating beam despite the many types of deformations caused by photon pressure and thermal expansion. There is also a requirement for structural stability, the ability to survive acceleration without collapse, and crumpling under acceleration, as depicted in the figure above. And there could be thermal and tensile failure as well as rupture of sail materials. Many studies of this issue have shown multiple solutions.
Stable designs exist for concave shapes and for flat flexible sails with millimeter scale photonic structures to control reflections. (Simple flat sails cannot achieve beam-riding stability because specular reflection produces forces only normal (perpendicular) to the surface.) A considerable advantage of flat sails is that curved sail shapes are more difficult to fabricate at meter scales. However, Starshot has shown that even flat sails can beam-ride by tailoring asymmetric optical properties to produce transverse restoring forces with millimeter-scale photonic structures to control reflections. So a flat sailcraft can be modified to scatter light as if it were curved. For example, the Swartzlander group, in a series of theoretical, computational, and experimental studies, has shown that a flat sail whose reflecting surface is equipped with diffractive gratings is directionally stable [8,9]. Anisotropic scattering of incident light into the grating diffraction orders manifests in optical restoring forces transverse to the membrane, redirecting incident photon momentum to produce beam-riding.
Such metagratings or metasurfaces consist of subwavelength scatterers shaped as disks, blocks, spheres, etc. shape the scattered wavefronts, redirecting incident photon momentum transversely. This provides stabilizing restoring forces and torques. However, adding metagratings makes the sail heavier than the ~0.1 gram per square meter goal. And photonic grating patterns would have to be produced over a large area. The advantage of flat sails will significantly streamline and simplify the fabrication process. The issue is whether such structures can be scaled to manufacture on the size of meters with low mass.
Spin-stabilization will likely be needed to prevent the collapse of sails while acceleration is underway. A beam can carry angular momentum and communicate it to a sail to help control it in flight. Spin can be modified remotely by circularly polarized beams from the ground [10]. It also allows ‘hands-off’ unfurling deployment through control of the sail spin at a distance [10-12]. Spinning them at ~100 Hz rates gyroscopically stabilizes sails against drift, yaw and tilting, allowing numerous shapes to retain their stability. (Circularly polarized electromagnetic fields carry both linear and angular momentum, which acts to produce a torque through an effective moment arm of a wavelength, so longer wavelengths are more efficient in producing spin.)
A final and crucial issue: Can the data be returned from distant space targets at sufficient data rates before the sail moves far beyond the star? For solar system-scale missions this is possible with existing microwave communication technologies. that were realized 50 years ago in the Deep Space Network. For interstellar missions it is possible by using laser communications. Though today’s laser communication systems are far too heavy for Starshot, which instead aims to operate part of its sail as an optical phased array. There are methods of making this likely in future decades [13]. That is because we understand essentially completely the fundamental limits on communication, and our technology today is able to operate very close to those limits.
The mission objective is to return 100 kB of data. The power requirement on board is driven primarily by the communication needs as well as pointing, tracking and computation. The energy technology is a thin film, radioisotope thermoelectric generator.
Propulsion-oriented scientists usually assume that the mission should be done at maximum speed. But information scientists’ relation to speed is different; they focus on how it affects the data return:
* Slower is better since observations are easier and there is more time in the vicinity of the target star.
* The measure of mission performance is the volume of data returned reliably vs the ‘data latency’ (defined as time from acquisition at Centauri to return to Earth of an entire observational data set).
So from this perspective speed is a secondary parameter except as it influences the data volume and data latency, which will relate to the payload mass, and in particular the communications mass.
Messerschmitt, Lubin and Morrison have studied the minimum data latency that can be achieved for a given data volume, or equivalently the maximum data volume that can be achieved for a given data latency [13, 14]. Generally, they reduce speed for high latency (with the benefit of larger data volume, so larger mass, more instrumentation, and larger data volume).
From this, the key insight that governs the difficult problem of returning data over interstellar distance is that a cost-optimized (meaning cost minimized) system scales as the relation between speed v and mass m: v~1/m1/4. That means we can have a much heavier communication system onboard. Achieving the data return is more credible. This leads to an optimum mass that maximizes data volume for a given data latency. Future communications research will deal with several probes downlinking concurrently from the same target star. Separating these downlinks (‘multiplexing’, using different formats, polarization, etc.) is very challenging,
That leads to a very significant development conclusion: We would of course develop heavier, lower velocity probes early on as the Beamer is being built out. The Beamer will be built by adding modules of power and aperture over time. It is likely what will happen is that technologies advance, such as sail materials are improved and mass is reduced. As faster solar system deep space missions occur, mass will either drop as the system performance improves or will increase for faster, better data return. That’s the natural development path, leading to faster, better missions.
The on-board pointing system of the sail is also a technical challenge. It must point in the direction of our Solar System, and the beam will be larger than Earth’s orbital diameter, 2 AU. That means a pointing accuracy of a milliarcsecond, about 10 microradians.
Phase I confirmed that short wavelength optical communications can provide the required down-link capability with limited data rate. Low-cost receiver aperture concepts were developed.
System Cost
Before I joined Starshot, I developed an analysis for cost optimization of beam-driven sail systems. In it, the trade-off was between the cost of the sources powering the array versus the cost of the array itself. That was in agreement with the cost of transmitter systems that had been built for interplanetary communications. My conclusion was that the minimum capital cost is achieved when the cost is equally divided between the array antenna and the radiated power [15].
However, Starshot requires more power than can be directly supplied by the normal electrical grid. Therefore, energy storage for the system has to be included, and becomes a substantial cost element [16, 17]. That results in a considerable change in the laser aperture, laser power, and energy storage cost. The result is that the laser cost, which is ~80% of the array cost, becomes the dominant element in the total project cost. The cost trends shown below demonstrate that cost is viable for future fiber amplifiers at ~$0.10/W, and future semiconductor lasers at ~$0.01/W.
The figure below shows that current laser fiber amplifiers and semiconductor laser costs are far too high to afford a Starshot system today. The hope is that economies of scale in the application of lasers to aspects of modern life, for example self-driving cars, will drive down the cost of lasers by economies of scale. In order to reach an affordable level for Starshot, the prices have to fall to order of cents per watt, not many dollars per watt we have today. The points at 2040 and 2050 shows what will have to occur if the cost of Starshot is to be of order 10 billion dollars. That requirement is two to three orders of magnitude cost reduction.
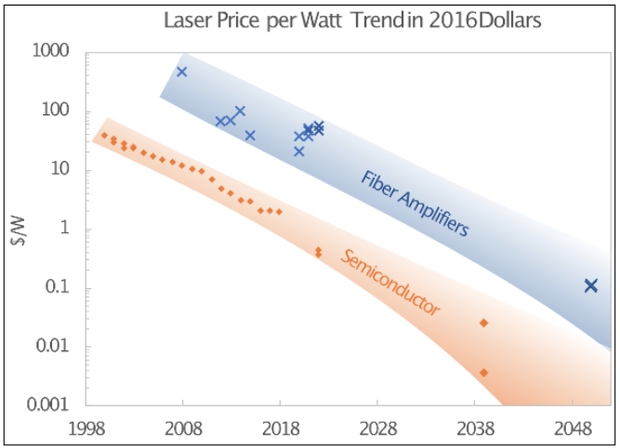
Image: Cost trends for fiber amplifiers and semiconductor lasers.
The Future of Beam-driven Sails
Phase II technical demonstrations, such as laboratory beam-riding sail flights and including orbital sail deployment and sail acceleration, would lead to a firm experimental basis for pilot production of the key sub-systems, leading to the beginning of array construction. That would later lead to precursor missions.
While the Beamer is under construction, many missions become possible that are at speeds lower than interstellar, as well as other applications. The laser driver can beam power to locations in space, such as Earth satellites and space stations. It can deorbit orbital debris. It can drive fast sail missions to the Moon, Mars and the outer planets. At Mars, it could have a second laser array to decelerate the spacecraft, or a retro reflector system, such as proposed by Forward, could reflect a beam from Earth to slow the sailcraft at Mars. And it can beam power to high-performance ion engines.
Development of fast sailcraft that can travel beyond our solar system will enable us to understand the interstellar medium and then, in the fast encounter with other star systems, acquire imaging, spectroscopy, and in situ particle and field measurements.
Beam-driven sails are the only way that probes can be sent to the stars in this century. Completion of Phase II would bring much-increased credibility to the concept by demonstrating beam-riding and operation of a Beamer module in the laboratory. Then the dream of beam-driven interstellar travel could be realized.
Kevin Parkin has even envisioned human beam-driven fast travel to the stars. Accelerating at Earth gravity to relativistic speeds, allowing us to contemplate human travel in future. He points out that human civilizations’ energy production doubled every 40 years since 1800, so that the energies needed for the simplest such missions will be attainable by the end of the century.
Acknowledgements: Figures are by permission of Breakthrough Starshot and Michael Kelzenberg. I also want to thank Kevin Parkin, Dave Messerschmitt and Al Jackson for technical discussions about Starshot.
References
1) Atwater, H. Starshot: from science to spacecraft to missions, Harry Atwater, Interstellar Research Group , Montreal 2023, https://www.youtube.com/watch?v=jV2sNOYzaFA
2) See also same title, Breakthrough Discuss, Harry Atwater, 2023 https://www.youtube.com/watch?v=IrLcllx0LpQ
3. Laser Propulsion in Space: Fundamentals, Technology, and Future Missions, Claude Phipps, ed., Elsevier., Cambridge, MA ,2024.
4. Worden S., Green, W. Schalkwyk, J., Parkin K., and Fugate R., “Progress on the Starshot Laser Propulsion System,” Applied Optics, doi: 10.1364/AO.435858, 2021.
5. Bandutunga C., Sibley P., Ireland M. J., and Ward, R., “Photonic solution to phase sensing and control for light-based interstellar propulsion”, J. Opt. Soc. of Am. B, 38, 1477-1486, 2021.
6. Benford, G., Goronostavea, O., and Benford, J., “Experimental tests of beam-riding sail dynamics” in Beamed Energy Propulsion, AIP Conference Proceedings 664, Pakhomov, A., Ed. 325, 2003.
7. Gao, R., Kelzenberg M. D., and Atwater H. A., “Dynamically Stable Radiation Pressure Propulsion of Flexible Lightsails for Interstellar Exploration”, Nature Comun, 15, 4203. https://doi.org/10.1038/s41467-024-47476-1, 2024,
8. Srivastava P., Chu Y., and Swartzlander G., “Stable diffractive beam rider,” Opt. Lett. 44, 3082-3085, 2019.
9. Chu Y., Tabiryan N. and Swartzlander G., Experimental Verification of a Bigrating Beam Rider. Phys Rev Lett. (123(24), 2024.
10. Benford, G., Goronostavea, O., and Benford, J., “Spin of microwave propelled sails,” in Beamed Energy Propulsion, AIP Conference Proceedings 664, Pakhomov, A., Ed., 313, 2003.
11. Benford, J. and Benford, G., “Elastic, electrostatic and spin deployment of ultralight sails”, JBIS 59 76, 2006.
12. Martin, P. et al., “Detection of a Spinning Object Using Light’s Orbital Angular Momentum” Science 341 537, 2013.
13. Messerschmitt D., Lubin P. and Morrison I., “Challenges in Scientific Data Communication from Low-mass Interstellar Probes”, ApJS 249,36, 2020.
14. Messerschmitt D., Lubin P. and Morrison I., “Interstellar flyby scientific data downlink design,” arXiv preprint arXiv:2306.13550, 2023.
15. Benford, J., “Starship Sails Propelled by Cost-Optimized Directed Energy”, JBIS 66, 85, 2013)
16 Parkin, K., “The Breakthrough Starshot Systems Model”, Acta Astronautica 152, 370–384, 2018.
17. Parkin, K., “Starshot System Model” in Laser Propulsion in Space: Fundamentals, Technology, and Future Missions, Claude Phipps, ed., Elsevier., Cambridge, MA ,2024.
Starshot Is a Success: Part I
The fortunes of Breakthrough Starshot have been the subject of so much discussion not only in comments in these pages but in backchannel emails that it is with relief that I turn to Jim Benford’s analysis of a project that has done significant work on interstellar travel and is still very much alive. Jim led the sail team for several of his eight years with Breakthrough Starshot and was with the project from the beginning. In this article and a second that will run in a few days, he explains how and why press coverage of the effort has been erroneous, and not always through the fault of writers working the story. Let’s now take a look at what Starshot has accomplished during its intensive Phase I.
by James Benford
“Make no mistake — interstellar travel will always be difficult and expensive, but it can no longer be considered impossible.” – Robert Forward
Breakthrough Starshot has not failed, nor has it been canceled. Phase I of the program achieved its stated objectives: to identify potential show-stoppers in beam-driven interstellar propulsion and determine whether credible solutions exist. That goal was met.
Recent media coverage, including a Scientific American cover article titled “Voyage to Nowhere,” misunderstands both the intent and the outcome of Phase I. The reality is that the project thus far has been successful. It was put “on hold, paused” in 2024 to restructure for the next phase and seek broader support. It has not been canceled, as some in the media are saying.
I contend that Starshot succeeded because the key Phase I objectives were met. Of course, extensive future effort in the later phases is needed to create a fully functional Starshot system, principally the beamer and sailcraft (referred in the project as “photon engine” and “lightsail”). The major issues have been found to have credible solutions. A great many Starshot-related papers have been published. Many address the crucial issues of sail materials and sail ‘beam-riding’, meaning staying on the beam while undergoing inevitable perturbations. There is a final report, but it has not yet been published.
The principal issues for Starshot were 1) Can a phased array of lasers be constructed that is sufficiently coherent and directive as well as being affordable? 2) Can a sail material be made that will have high reflectivity, very low absorption, high emissivity and very low mass so as to be efficiently accelerated and not overheat? 3) Can a sail ride stably on the beam because of inherent restoring forces (without feedback, which is impossible over long ranges)? 4) Can data be sent back to Earth from the probe at sufficient data rates before the sail moves far beyond the target star?
In this first of two reports on the successes of the Starshot project, I discuss the shape of later phases in the effort, and distortions in the reporting on it. In the second report I will describe the major accomplishments of Phase I.
Starshot was not initiated to fully design, build and launch the first interstellar ‘lightsail’ (as they are called, referring to both the low mass and the near-visible frequency of the laser). The program path was divided into phases, as shown below. The first phase was to invest in high-risk, high-reward research that would de-risk the technology. Phase 1 was to find if there were any ‘show-stoppers’ and pave the way forward. It accomplished that.
High levels of research by Starshot retired most of these key issues for beam-driven sail systems, at least at the conceptual level. The results are at the TRL 2 level. Experiments are needed to verify the solutions for these major issues found in Phase 1.
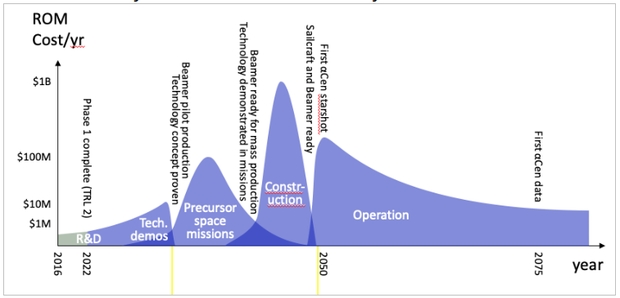
In Phase II, a coalition of Caltech and other institutions would lead experimental technical demonstrations, and the first experiments in orbit. Then, with the technology concepts having been proven, it’s on to near-term missions shaking out various technologies while performing precursor missions, probably to the outer solar system. Much effort would be needed in systems engineering to enable such precursor missions.
The first phases of Starshot, the R&D program, are projected to cost $120M, which includes Phase 1, and concludes with solar system science missions in the medium-term. The large effort would then follow: construction of the Starshot System and finally, operation of the System and the first interstellar probe voyages.
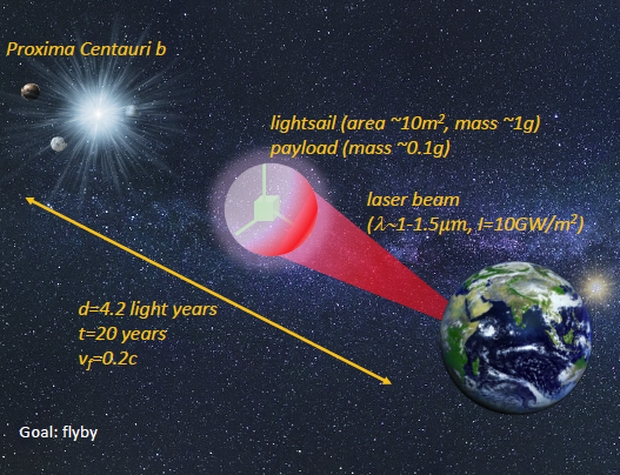
Many requirements of the Starshot mission come together at the sail. Principal technical issues are the design of the beamer, material to be used and whether the beam and sail stay together, meaning stable beam-riding by the sail:
• Stability is influenced by sail shape, beam shape and the distribution of mass, such as payload, on the sail.
• Material properties, are its reflectivity, absorptivity and transmissivity, it’s tensile strength and its areal mass density.
• Deployment of the diaphanous sail, correctly oriented and including any initial spin, is of course a key requirement.
• The beamer interacts with the sail through its power distribution on the sail-causing differential stresses. This depends on duration of the acceleration, the transverse width of the beam, pointing error of the beam as well as its pointing jitter.
• Data return to Earth, interstellar communications, is perhaps the greatest challenge of all.
What Scientific American got wrong
Journalism is only the first draft of history, so flaws occur. Assessing a system as complex as Starshot is a challenge to a journalist with limited time. It would take years to read and absorb all the relevant literature and to mentally organize it into a reconciled and coherent understanding of the system as a whole.
The biased title – “Voyage To Nowhere” – of the piece in Scientific American, (which was chosen by the editors, not the author Sarah Scoles), may have been chosen to refer to the famous Bridge to Nowhere in Alaska and the Train to Nowhere in California. The Scientific American reporting is already being mistaken for a primary source by others, who are stating that Starshot has been “canceled”. This is an example of how media myths, once manufactured, propagate through journalistic copying.
The article fails to understand the Starshot project for a basic reason: The key people who did extensive work on the program were not available or not even known to the writer.
Because the principal workers from the Breakthrough Foundation and the leaders of Breakthrough Starshot, Pete Worden and Avi Loeb, were not interviewed, it seems the author did not know who the main contributors actually were. She relied instead on people she could easily reach. Few of them are major contributors to the program and most left the project early on or never actually participated in the project. A key participant who is not mentioned is Kevin Parkin [1, 2], who spent 8 years under contract, as did most of us who were in at the beginning or even before that. Others are Mason Peck (who is mentioned in the piece), Paul Mauskopf and Dave Messerschmitt. Unfortunately, the final report, which went through many iterations, has never been published publicly [3].
The recent policy of Breakthrough Starshot has been to have little contact with the media, so not to engage with Sarah Scoles at all didn’t help things: it left the door open for detractors to influence the narrative in her piece. Communication was a priority, with public outreach from and within Starshot during Phase 1. In research, communication enables cross-fertilization and prevents work duplication. The big gap now is a comprehensive publication that ties it all together. It could motivate researchers to continue or take up the project later if Phase II occurs.
The article also truncates the long history that led up to Starshot. Beam-driven propulsion concepts didn’t start in 2016! This was documented in my Photon Beam Propulsion Timeline, which appeared here at the start of Starshot in 2016. Media are not aware of how much has been done by the propulsion community over the last decades. Several areas of photon beam-driven sail system development, to include experiments demonstrating sail beam-driven flight [4, 5] and sail stability and dynamics, such as beam-driven spin of sails for stability [6, 7], have been reserched. The major innovation which caused the beginning of Starshot was the realization that going to much smaller sails and much higher accelerations reduces the cost of the overall system substantially.
The budget estimate given in the Scientific American article is clearly wrong. That only 4.5 million dollars could fund 8 years of steady work by many people is absurd. Thirty contracts were executed over 8 years. There were years of invitational meetings, a standing staff of advisors, subcommittees for specific topics; all of them further expenditures. And I count about 50 Starshot-related papers, some of which have been published since it was put on hold. I estimate that Breakthrough Starshot Phase 1 had a cost of 25 million dollars.
The way Forward
Phase II would lead to a firm experimental basis for the later phases in Figure 1. If Breakthrough decides to move on to Phase II, it must deal with the costs of interruption: institutional knowledge about the previous work, which is never fully captured in documentation, will need to be relearned, as the people who worked on Phase 1 have dispersed to other programs.
My second piece on Breakthrough Starshot, scheduled to run here next week, will describe the present state of the concept and the many advances achieved by Starshot in Phase I
Breakthrough Starshot was the most significant event in the history of beam propulsion, which clearly is the only way that probes can be sent to the stars in this century. And now the work goes on, the hope still lives, and the dream of beam-driven interstellar travel could be realized.
References
[1] “The Breakthrough Starshot Systems Model”, Kevin Parkin, Acta Astronautica 152, pp 370–384 (2018).
[2] “Starshot System Model” Kevin Parkin, Ch 3, in Claude Phipps, Editor, Laser Propulsion in Space: Fundamentals, Technology, and Future Missions, Elsevier (2024).
[3] Breakthrough Starshot Summary Report, September 2023, not published.
[4] “Microwave Beam-Driven Sail Flight Experiments”, James Benford, Gregory Benford, Keith Goodfellow, Raul Perez, Henry Harris, and Timothy Knowles, Proc. Space Technology and Applications International Forum, Space Exploration Technology Conf, AIP Conf. Proceedings 552, ISBN 1-56396-980-7STAIF, pg. 540, (2001).
[5] “Laser-Boosted Light Sail Experiments with the 150 kW LHMEL II CO2 Laser,” Leik Myrabo, Timothy Knowles, John Bagford and H. Harris, “High-Power Laser Ablation IV,” edited by Claude Phipps, Editor, Proc. Space Exploration Technology Conf., 4760 pp. 774-798 (2002).
[6] “Spin of Microwave Propelled Sails” Gregory Benford, Olga Goronostavea and James Benford, Beamed Energy Propulsion, AIP Conf. Proc. 664, pg. 313, A. Pakhomov, ed., (2003).
[7] “Experimental Tests of Beam-Riding Sail Dynamics”, James Benford, Gregory Benford, Olga Gornostaeva, Eusebio Garate, Michael Anderson, Alan Prichard, and Henry Harris, Proc. Space Technology and Applications International Forum (STAIF-2002), Space Exploration Technology Conf, AIP Conf. Proc. 608, ISBN 0-7354-0052-0, pg. 457, (2002).
The Language of Contact
How we think intersects with the language we think in. Consider the verb in classical Greek, a linguistic tool so complex that it surely allows shadings of thought that are the stuff of finely tuned philosophy. But are the thoughts in our texts genuinely capable of translation? Every now and then I get a glimpse of something integral that just can’t come across in another tongue.
Back in college (and this was a long time ago), I struggled with Greek from the age of Herodotus and then, in the following semester, moved into Homer, whose language was from maybe 300 years earlier. The Odyssey, our text for that semester, is loaded with repetitive phrases – called Homeric epithets – that are memory anchors for the performance of these epics, which were delivered before large crowds by rhapsōdoi (“song-stitchers”). I was never all that great in Homeric Greek, but I do remember getting so familiar with these ‘anchors’ that I was able now and then to read a sequence of five or six lines without a dictionary. But that was a rare event and I never got much better.
The experience convinced me that translation must always be no more than an approximation. A good translation conveys the thought, but the ineffable qualities of individual languages impose their own patina on the words. ‘Wine-dark sea’ is a lovely phrase in English, but when Homer spins it out in Greek, the phrase conjures different feelings within me, and I realize that the more we learn a language, the more we begin to think like its speakers.
My question then as now is how far can we take this? And moving into SETI realms, how much could we learn if we were actually to encounter alien speakers? Is there a possibility of so capturing their language that we could actually begin to think like them?
Let’s talk about Ted Chiang’s wonderful “Story of Your Life,” which was made (and somewhat changed) into the movie Arrival. Here linguist Louise Banks describes to her daughter her work on aliens called heptapods, seven-limbed creatures who are newly arrived on Earth, motives unknown, although they are communicating. Louise goes to work on Heptapod A and Heptapod B, the spoken and written language of the aliens respectively.

Image: A still from Denis Villeneuve’s 2016 film Arrival captures the mystery of deciphering an alien language.
Heptapod B is graphical, and it begins to become apparent that its symbols (semagrams), are put together into montages that represent complete thoughts or events. The aliens appear to experience time in a non-linear way. How can humans relate to that? Strikingly, immersion in this language has powerful effects on those learning it, as Louise explains in the story:
Before I learned how to think in Heptapod B, my memories grew like a column of cigarette ash, laid down by the infinitesimal sliver of combustion that was my consciousness, marking the sequential present. After I learned Heptapod B, new memories fell into place like gigantic blocks , each one measuring years in duration, and though they didn’t arrive in order or land continuously, they soon composed a period of five decades. It is the period during which I knew Heptapod B well enough to think in it, starting during my interviews with Flapper and Raspberry and ending with my death.
Flapper and Raspberry are the human team’s names for the two heptapods they’re dealing with, and we learn that Louise now has ‘memories’ that extend forward as well as back. Or as she goes on to explain:
Usually, Heptapod B affects just my memory; my consciousness crawls along as it did before, a glowing sliver crawling forward in time, the difference being that the ash of memory lies ahead as well as behind: there is no real combustion. But occasionally I have glimpses when Heptapod B truly reigns, and I experience past and future all at once; my consciousness becomes a half-century long ember burning outside time. I perceive – during those glimpses – that entire epoch as a simultaneity. It’s a period encompassing the rest of my life, and the entirety of yours.
The ‘yours’ refers to Louise’s daughter, and the heartbreak of the story is the vision forward. What would you do if you could indeed glimpse the future and see everything that awaited you, even the death of your only child? How would you behave where your consciousness is now, with that child merely a hoped for future being? How would such knowledge, soaked in the surety of the very language you thought in, affect the things you are going to do tomorrow?
A new paper out of Publications of the National Academy of Sciences has been the trigger for these reflections on Chiang’s tale, which I consider among the finest short stories in science fiction history. The paper, with Christian Bentz (Saarland University) as lead author, looks at 40,000 year old artifacts, all of them bearing sequences of geometric signs that had been engraved by early hunter-gatherers in the Aurignacian culture, the first Homo sapiens in central Europe. It was a time of migrations and shifting populations that would have included encounters with the existing Neanderthals.
These hunter-gatherers have left many traces, among which are these fragments that include several thousand geometric signs. What struck me was that these ancient artifacts demonstrate the same complexity as proto-cuneiform script from roughly 3000 BC. Working with Ewa Dutkiewicz (Museum of Prehistory and Early History of the National Museums, Berlin), Bentz notes objects like the ‘Adorant,’ an ivory plaque showing a creature that is half man, half lion. Found in the “Geißenklösterle,” a cave in the Achtal Valley in southern Germany, it’s marked by notches and rows of dots, in much the same way as a carved mammoth tusk from a cave in the Swabian Alb. The researchers see these markings as an early alternative to writing. Says Bentz:
“Our analyses allow us to demonstrate that the sequences of symbols have nothing in common with our modern writing system, which represents spoken languages and has a high information density. On the archaeological finds, however, we have symbols that repeat very frequently – cross, cross, cross, line, line, line – spoken languages do not exhibit these repetitive structures. But our results also show that the hunter-gatherers of the Paleolithic era developed a symbol system with a statistically comparable information density to the earliest proto-cuneiform tablets from ancient Mesopotamia – a full 40,000 years later. The sequences of symbols in proto-cuneiform are equally repetitive; the individual symbols are repeated with comparable frequency. The sequences are comparable in their complexity.”

Image: The so-called “Adorant” from the Geißenklösterle Cave is approximately 40,000 years old. It is a small ivory plaque with an anthropomorphic figure and several rows of notches and dots. The arrangement of these markings suggests a notational system, particularly the rows of dots on the back of the plaque. Credit: © Landesmuseum Württemberg / Hendrik Zwietasch, CC BY 4.0.
As the researchers comment, the result is surprising because you would think early cuneiform would be much closer in structure to modern systems of notation, but here we have, over a period of almost 40,000 years, evidence that such writing changed little since the Paleolithic. Says Bentz: “After that, around 5,000 years ago, a new system emerged relatively suddenly, representing spoken language—and there, of course, we find completely different statistical properties,”
The paper digs into the team’s computer analysis of the Paleolithic symbols, weighing the expression of information there against cuneiform and modern writing as well. It’s clear from the results that humans have been able to encode information into signs and symbols for many millennia, with writing as we know it being one growth from many earlier forms of encoding and sign systems.
We have no extraterrestrials to interrogate, but even with our own species, we have to ask what the experience of people who lived in the Stone Age was like. What were they trying to convey with their complex sequences of symbols? The authors assume they were as cognitively capable as modern humans and I see no reason to doubt that, but how we extract their thought from such symbols remains a mystery to be resolved by future work in archaeology and linguistics.
And I wonder whether Ted Chiang’s story doesn’t tell us something about the experience of going beyond translation into total immersion in an unknown text. How does that change us? Acquiring a new language, even a modern one, subtly changes thought, and I’m also reminded of my mother’s Alzheimer’s, which somehow left her able to acquire Spanish phrases even as she lost the ability to speak in English. I always read to her, and when I tried to teach her some basic Spanish, the experiment was startlingly successful. That attempt left me wondering what parts of the human brain may be affected by full immersion in the language of any future extraterrestrial who may become known to us.
Ludwig Wittgenstein argued that words only map a deeper reality, saying “The limits of my language mean the limits of my world.” Beyond this map, how do we proceed? Perhaps one day SETI will succeed and we will explore that terrain.
The paper is Bentz et al., “Humans 40,000 y ago developed a system of conventional signs,” Publications of the National Academy of Sciences 123 (9) e2520385123. 23 February 2026. Full text. For more on this work, see Bentz and Dutkiewicz’ YouTube video: https://www.youtube.com/@StoneAgeSigns. Thanks to my ever reliable friend Antonio Tavani for sending me information about this paper.

