Claudio Flores Martinez has just finished an MSc at the European Molecular Biology Laboratory (EMBL) in Heidelberg, Germany and is now enroute to a PhD in theoretical biology. He currently serves as a research assistant at the Developmental Biology Unit of EMBL and the University of Heidelberg’s Centre for Organismal Studies. With three papers in Acta Astronautica under his belt, Claudio is already deep into theoretical evolutionary biology, and in particular the contingency vs. convergence debate. In the paper below, he discusses how these issues couple with the possible emergence and development of life on Europa and the potential biosignatures by which we might find it, a journey that takes us deep into the nature of living systems. Just what might we encounter under that Europan ice?
by Claudio Flores Martinez

“Science is an endless search for truth. Any representation of reality we develop can be only partial. There is no finality, sometimes no single best representation. There is only deeper understanding, more revealing and enveloping representations. Scientific advance, then, is a succession of newer representations superseding older ones, either because an older one has run its course and is no longer a reliable guide for a field or because the newer one is more powerful, encompassing, and productive than its predecessor(s).”
— Carl R. Woese, “A New Biology for A New Century” (2004) [1]
A useful Gedankenexperiment to ponder the implications of a putative alternate emergence of life on Jupiter’s moon Europa, is to adopt the perspective of a future field astrobiologist who finds himself submerged in the abyssal depths of an alien ocean, employing the technology of a far future embodiment of planetary bathy-nautics. Not relegating the complex process of biosignature detection to the highly advanced sensory equipment attached to his exosuit, we might wonder what it actually is that he sees beyond the other side of his metamaterial helmet visor? What is he going to observe as he is slowly drifting through the vitellus of the precious world egg mankind came to call Europa? Impelled to turn off the on-board lights by an inner voice, a transcendental moment of perfect stillness commences. Nothing moves out there.
Eternity, however, has not been reached yet. Our bathy-naut, let’s call him Castor for the present purpose, is still breathing, taking in the splendour of pure discovery. He has been trained for this task all along, but he is now checking his vital signs as he believes himself to be deceived by his sensory-deprived brain or some inorganic deep-sea type of ignes fatui – – – pinpoints of light are flaring up all around him as he is moving the arms of his mechanical exo-skeleton around in amazement. From the distance a bluish, fragile jelly-like creature is approaching, cloaked in sapphirine light. The luminosity of the Europan life form is increasing as it is getting closer, reminding him of the intense glow of the fusion plasma that accompanied their journey to the Outer System and, ultimately, brought them here. Reaching out with his right glove, cast in a pressure-proof titanium alloy, his mind is electrified by one thought alone: man – finally – has been handed the Promethean light – not in the form of fusion technology as so many have believed – but, instead, it is incarnate by the organic emanation of spectral blue bioluminescence…
Image: Is there a light shining in the eternally sunless ocean of Europa after all? Credit: JPL/NASA/Steve Burg (overlay of the composite “Galileo” imagery and 1988 artwork for James Cameron’s “The Abyss”)
INTRODUCTION
In the light of a potential discovery of extraterrestrial life through advanced robotic exploration (most likely on the icy moons of the outer Solar System, Europa and Enceladus) or astronomical observations, a future theory of universal biology, if it exists, should be able to make falsifiable predictions about the kind of biological activity such missions expect to encounter. To do so, current evolutionary thinking has to provide a comprehensive explanation of the convergent emergence of numerous adaptive traits that evolved independently and repeatedly across and within all three domains that make up the tree of life. Prime examples include distinct cellularization events in eubacteria and archaea, endosymbiosis, multicellularity, bioluminescence, echolocation, fusiform shapes and intelligent and social behavior in nonprimate species.
The long-term exploration strategy of Europa and its potential global subsurface ocean envisions the future in situ probing for life of the local aquatic environment with an integrated approach that uses a melt probe carrying a second-stage autonomous underwater vehicle (AUV). However, accurately predicting the exact nature of putative biological activity on Europa is extremely difficult. This is due to an unsolved and fundamental problem in theoretical evolutionary biology, namely the contingency vs. convergence debate. It is far from certain if stable trajectories exist, leading from mechanistically identical origins of life towards higher forms of biological organization, which are independently and repeatedly, and thus in a convergent manner, traced by the process of evolution within planetary biospheres apart from Earth. Therefore it appears to be helpful, first on theoretical grounds, to address the possibility of convergent evolution in two unrelated evolutionary systems that emerged and subsequently evolved and developed independently.
Recent studies indicate that convergent evolution is actually occurring on the molecular sequence level and, indeed, genome-wide in some instances. Information and (biological) systems theory are beginning to reframe the phenomenon of convergent evolution in terms of top-down causation and resulting functional equivalence classes. One instantiation of convergent evolution, bioluminescence, is especially well-suited for studying its underlying informational, genomic, functional and ecological basis. In addition, as we will see, bioluminescence is a promising candidate in terms of predictable higher-order biocomplexity that might have evolved in parallel on both Earth and Europa. Research on living light within terrestrial oceans can lay the theoretical and practical foundation for a dedicated agenda on defining potential biosignatures encountered within Europa’s subsurface ocean. Exploring the wonders of bioluminescence on the surface and in the depths of Earth’s oceans will facilitate the technology development of adequate robotic infrastructure that is needed to realize the dream of navigating an alien ocean.
COSMIC CONVERGENT EVOLUTION
Can science pin down universal attractors – on the level of biochemistry, function and morphology or even on ecological and cultural scales – within the unknown hyperspace or astrobiological landscape of biological potential? This fundamental question transcends the boundaries of any particular biological discipline and takes the issue of extraterrestrial evolution into the realms of philosophy, complexity theory and cosmology, opening up the way for an emerging scientific view of the Universe in which life is a natural and repeated cosmic phenomenon. The detection of a genuine second origin of life within our Solar System, imaginably on Jupiter?s icy moon Europa, or through a future success of the SETI enterprise could pave humanity?s way into a new era of universal scientific insight and recast its lonesome role within the cosmic scenery [2]. Far from being a contingent “cosmic accident”, as has been influentially suggested previously (for example by pioneer of molecular biology Jacques Monod and paleontologist Stephen J. Gould) life on Earth in general and Homo sapiens in particular can be reconceived of as natural and necessary outcomes of cosmic evolution, a process that is spanning almost infinite expanses of space and time, while, as will be argued here, being an inherent function, an adaptation of the living Universe, the Biocosm (a term taken from the magnificent work of James N. Gardner).
I am part of an international group of scientists, the Evo Devo Universe (EDU) research community, founded by systems theorist John Smart and philosopher Clément Vidal in 2008. We are seeking to envision, elaborate, discuss, test and promote a guiding vision for the emergence of a 21st century universal science of complexity that is able to reconcile thermodynamics with the onset of replication in the prebiotic realm, and to better understand the major transitions in evolution – from protocells to integrated nervous systems in multicellular organisms – as a necessary outcome of a cascading unfoldment, a process of universal complexification, and evolutionary development. Such processes may find their origin in the fundamental physical make-up of our Cosmos, the fabric of the observable Universe, whose threads, the laws of Nature, its DNA, we are beginning to unravel by means of our imagination, rational mind, simulation, and experimentation. Many researchers think of physics as the foundation for chemistry and biology. For the longest part of the past century or two this has been the case indeed. Now, however, we are getting first glimpses of the true vastness of networked biological complexity encapsulated in a single bacterial cell, exponentiated in multicellular life forms, staggering in the workings of our own minds, and, finally, transcended with the imminent advent, for better or worse, of autonomously reproducing and evolving technological replicators:
“The underlying paradigm for cosmology is theoretical physics. It has helped us understand much about universal space, time, energy, and matter, but does not presently connect strongly to the emergence of information, computation, life and mind. Fortunately, recent developments in cosmology, theoretical biology, evolutionary developmental biology, information and computation theory, and the complexity sciences are providing complementary yet isolated ways to understand our universe within a broader ‘meta-Darwinian’ framework in which contingent and selectionist or “evolutionary” and convergent, hierarchical, and replicative or “developmental” processes appear to generate complexity at multiple scales. The rigor, relevancy, and limits of an “evolutionary developmental” approach to understanding universal complexity remains an open and understudied domain of scientific and philosophical inquiry.”
— EDU’s mission statement (www.evodevouniverse.com)
A new expanded theory of evolution appears to be a prerequisite for the successful development of astrobiology and the complementary SETI studies. Here it will be outlined under the term Cosmic Convergent Evolution (CCE). The latter proposition, which is based on a growing volume of related literature, is a powerful updated view of life and a synthesis of evolutionary theorizing within the disciplines of astrobiology and cosmology, which contends that natural selection and evolutionary development in unison are a universal force of nature that might potentially lead to the emergence of similarly adapted life forms in analogous planetary biospheres.
First, the Universe may be envisioned as an evolving and, consequently, developing system. It might even comprise a replication cycle in which adaptive processes of its biological components occur to facilitate the generation and increase proliferation of these respective elements of the system and, eventually, enable replication of the Universe as a whole. During this process of cosmic ontogenesis the complexity of the system?s constituents increases naturally, and life and intelligence might be of paramount importance in catalyzing and structuring the emergence of a cosmologically extended biosphere. Redundancy of core regulatory and functional elements is a hallmark of any information-based replication system. Therefore, the universal replicator, or Biocosm, organizes itself in a way as to give rise to the repeated emergence of life and very likely, although less frequently, also to the evolution of highly advanced intelligence.
Second, in the continuing process of emerging biological complexity interplanetary and -stellar convergence could lead to predictable configurations in the fundamental organization of living matter. Just as there are attractors in physical phase space, biological systems might be biased towards a set of definable regions within the theoretical hyperspace of complexity.
The driving force behind interplanetary convergence is an assumed natural propensity of our Universe towards biogenesis. In such a scenario life should be emerging naturally as a complex far from-equilibrium phenomenon within aqueous environments that sustain geochemical energy gradients and offer the chemical building blocks for life as we know it. Further, recent theoretical (and some empirical) studies are starting to go beyond the “life is chemistry” paradigm by exploring how basic properties of biological systems, such as self-assembly, -organization, and replication, could be derived and quantitatively described by using fundamental physical laws, governing the state of any thermodynamic disequilibrium present in the physical world. Conversely, it is convincingly being argued that the potentialities of biological complexity might actually be able to constrain a completely new kind of physics [3].
Two distinct origins of life on Earth and, for example, Europa (resulting from a very similar prebiotic context) are by definition not identical in their genetic origin, but both are necessarily products of the same universal process – cosmic evolution. In this sense, separate events of biogenesis constitute convergent adaptations of a cosmologically extended biosphere, guided by an as of yet undiscovered natural principle of biogenicity. Just as the evolution of different species on Earth in environments which expose organisms to selective pressures of the same kind leads to the emergence of similarly adapted life forms, convergence across analogous planetary niches might result in predictable patterns of biological complexity. Adaptations of terrestrial organisms which repeatedly have been found to be convergent in nature, ranging from multicellularity to non-human intelligence, might be indicative of the robust evolution of comparable features in extraterrestrial life as well.
Through the ages speculative minds, often in mythological and sometimes in more naturalistic terms, have contemplated the idea of our own Universe being an organismic system. In the following I will argue that this idea is more than a mere theorem or speculation and, operating at the very forefront of scientific enquiry, is actually presenting us with falsifiable hypotheses that can be put to the test within this century. The possibility of a repeated and independent origin of life can be adequately described as a systemic property of a universal replicator that possesses a developmental cycle. Biological life proliferates at the onset of this sequence but might not prevail during later stages, which would be driven by a dramatic increase in cultural and technological complexity. Artificial forms of life and intelligence could be more apt in completing the cycle. A growing number of futurists are arguing that humanity’s developmental transition will occur within the 21st century and that a “Singularity”-type event will prove the reality of an Evo Devo Universe or Biocosm (consider for example John Smart’s daringly innovative “Transcension Hypothesis” [4]).
Personally, however, I am more inclined to view the Promethean plight fulfilled by clearly establishing the organic convergence between life on Earth and another celestial body. Ultimately, as the truly open-minded scientists that we profess to be, we should consider – tentatively – what a continued failure of robotic and astronomical astrobiological exploration within the Solar System and beyond, as well as an absent merger with the machine in the future, would imply about the nature of the reality we find ourselves in.
Ending this philosophical excursion on an optimistic note I would like to remind the readers of the introductory quote by Carl R. Woese, whom I consider to be one of the greatest but least known pioneers of biology from the 20th century (NASA established the “Institute for Universal Biology” in his honor). I herewith humbly submit that “Cosmic Convergent Evolution” might be a “more revealing and enveloping representation” for the emergence, evolution and development of biological complexity on Earth and, by extension, throughout the Cosmos.
ORIGIN OF LIFE, MINIMAL CONVERGENT TRAITS AND BIOSIGNATURES
In order to better understand how the most basic entity, which we could identify as a living organism, should look like, let us turn to the origin of biological evolution again. Life on Earth has evolved in a manner suggestive of a minimum set of prerequisites which comprises liquid water, biogenic elements (C, H, N, O, P, and S) and biologically usable energy (i.e. transducible into chemical bonds for storage and later usage in metabolic reactions). In regard to the most pressing constraint for the onset of a potential biosphere on Europa, for instance, namely available energy sources, recent studies including theoretical work and on-going hydrothermal laboratory experimentation have stressed the importance of evolutionarily conserved molecular “disequilibrium converting engines” in the proto-metabolism of early terrestrial life.
This work is part of the “Submarine Hydrothermal Alkaline Spring Theory” for the emergence of life [5]. It posits that off-ridge alkaline hydrothermal springs reacted with the metal-rich carbonic Hadean Ocean. During this reaction compounds such as hydrogen, methane and ammonia, as well as calcium and traces of acetate, molybdenum and tungsten were released by progressing serpentinization of ultramafic rock, thereby offering a sustained source of chemically transducible energy for early biological systems. Employing an ancient molecular apparatus resembling the ATP synthase complex, universally observed in extant life, to harvest the free energy contained in the resulting proton and redox gradients, these ancestral organisms could have been able to maintain and gradually increase the complexity of their metabolic and replicator systems. Assuming that conditions during the late stages of Europa?s planetary differentiation might have been comparable to Earth?s Hadean Ocean in terms of a rocky mantle exchanging chemicals and heat with an overlying slightly acidic water layer, many studies point at the importance of putative hydrothermal circulation systems as possible environments conducive to an alternative origin of life.
In analogy to the triad of parameters that is defining habitability for life as we know it, I introduced a framework for constraining the biocomplexity of putative alien biological systems [6]. Just as a given world must possess three basic requirements (liquid water, biogenic elements and an energy source) to be rendered habitable, biological entities that we could identify as such, should display three universal Minimal Convergent Traits (MCTs): 1) molecular replication and inheritance, 2) cellularization via membrane systems and 3) metabolic networks coupling energetically favorable biochemical reactions. These MCTs represent systemic properties of an alien organism. It is not implied, however, that extraterrestrial biology necessarily employs an identical set of molecules known from terrestrial life to sustain these features. In any case, if one of these traits is lacking in extraterrestrial organisms, we could not recognize them as life as we know it (and they would not be viable according to our current understanding of biology in the first place).
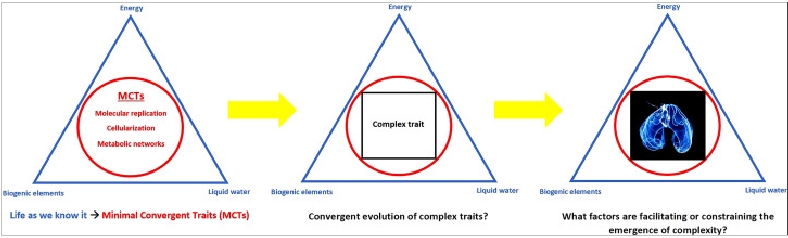
Image: Constraining putative alien biocomplexity by introducing Minimal Convergent Traits (MCTs): molecular replication, cellularization and metabolic networks. MCTs represent systemic properties rather than molecular identities. They delineate the phase space of potential higher-order evolutionary innovation on Europa. Figure from (Flores Martinez 2015) [6]
Research on the origin of life on Earth can give valuable insights into the principle mechanisms that molecule assemblies that instantiate MCTs have to be able to carry out, such as autocatalytic processes of self-replication. It is easier to reasonably predict complex or systemic properties of extraterrestrial life, like the previously mentioned characteristics of molecular replication, cellular organization and metabolic networks, than to anticipate the exact types of molecules involved in these functions. It is at the level of sufficiently advanced complexity of living systems at which the hypothesis of CCE proposes an evolutionary robustness of certain basic traits, like cellularization, and even higher evolved adaptations, for instance bioluminescence, among earthly and alien life. Generally, a universal mechanism which allows, first, the emergence of life on a given world and then, second, shapes its evolution according to commonly shared evolutionary trajectories toward increased biological complexity could be termed substrate-independent convergent evolution.
Such a definition does not imply that alien life can emerge from a more or less random variety of chemical elements. Almost certainly it would be based on carbon and organic molecules known from terrestrial biology. However, the specific substrates of the cellular replication and membrane components could be constituted by different biopolymers than those present in terrestrial organisms. One example for such a kind of convergence on Earth is the repeated emergence of cell membranes in archaea based on glycerol-ether lipids with isoprenoid sidechains rather than glycerol-ester lipids composed of fatty acid tails as found in eubacteria and eukarya.
Nevertheless, already on the level of proteins and their enzymatic functions it is difficult to imagine equally effective bio-catalytic molecular complexes. Whether molecular evolution on another world necessarily encompasses a hierarchical flow of genetic information starting from a replicating information-saving molecule (DNA) that gets transcribed into a messenger molecule (RNA) which in turn forms the basis for the eventual translation of the required cellular constituents, corresponding to terrestrial proteins, cannot be known with absolute certainty. For any biologist, on the other hand, it is hard to envision a similarly ingenious form of regulatory organization of genetic information, allowing multi-level tweaking of cellular processes. Given the integration of multiple molecule assemblies into functional networks in extraterrestrial cells, convergence of biological organization on a very profound level would become apparent.
According to NASA’s 2008 astrobiology roadmap a biosignature is defined as: “an object, substance and/or pattern whose origin specifically requires a biological agent. The usefulness of a biosignature is determined, not only by the probability of life creating it, but also by the improbability of nonbiological processes producing it. An example of such a biosignature might be complex organic molecules and/or structures whose formation is virtually unachievable in the absence of life. A potential biosignature is a feature that is consistent with biological processes…”. Future planetary exploration missions and spectroscopic exoplanet surveys will be limited due to a maximum volume available for scientific payload and highly sought-after observation time. Dedicated astrobiological space exploration is therefore dependent on a selection of previously chosen indicators for the presence of extraterrestrial life. More precisely, these indicators could be termed convergent biosignatures, as the mentioned instrumentation will scan for markers analogous to phenomena observed in terrestrial biota [7].
MCTs should suffice to delineate universal features of alien (microbial) life. More complex adaptations could be derived from this baseline by considering planetary properties of the search target. As already described, universal features of any kind of extraterrestrial life encompass: molecular replication, cellularization and metabolic networks. From these systemic properties more precise convergent biosignatures, i.e. potential molecular identities, can subsequently be deduced. Exactly this type of predictability is essential to the astrobiological endeavor, because future planetary exploration missions or space-telescope based exoplanet surveys need to be guided by a defined, or at least constrained, set of possible biosignatures. Here it will be argued that the notion of convergent evolution is pivotal in lending a certain degree of predictability to astrobiology and the related SETI field. Luckily, I have been already part of a pioneering lander mission design study for future in situ exploration of the plume fractures found on Enceladus [8]. The organizational scheme of MCTs adequately fits routine procedures in space mission design, namely the overarching science goal definition process via “science traceability matrices” (STMs) connecting science goal, science objectives, measurements, and detection methods in a logical and modular fashion.
| Science goal | Science objectives | Measurements | Detection methods |
|---|---|---|---|
| In situ probing of an aquiferous fracture to search for biosignatures | Molecular replication | Analyze water sample for presence of replicatory polynucleotides | Antibody microarray/sequencing; Nanopore-based instruments |
| Cellularization | Analyze water sample for presence of cellular structures and membrane material | Microscopy Flow cytometry Fatty acid markers Mass spectrometry |
|
| Metabolic networks | Analyze water sample for presence of expected metabolic products (CH4) | Mass spectrometry |
Science Traceability Matrix (STM) for a lander mission to Enceladus based on dedicated astrobiology science objectives (= MCTs). Instrument selection will be constrained by a logical and modular workflow according to the overarching science goal. Figure from (Konstantinidis et al. 2015) [8]
TOP-DOWN CAUSATION, INFORMATION HIERARCHIES AND MODULARITY
On an abstract systems theoretical level convergent evolution of organismal complexity is the result of biomolecular networks that are organized and structured by information hierarchies via top-down causation. The emergence of fundamental network properties such as modularity and the related phenomenon of functional equivalence classes of lower level operations or subroutines – both in biological and technological systems – can be explained as a corollary of top-down organized information hierarchies.
Top-down causation refers to the causal role of information in living systems. More specifically, it describes the process whereby “higher levels of organization in structural
hierarchies constrain the dynamics of lower levels of organization” [9]. Top-down theorists (e.g. Arizona State University physicist Sara I. Walker who is leading ASU’s “Emergence” group and has contributed a number of highly innovative and inspiring papers) propose that the organization, structure and function of living systems cannot be completely explained by a reductionist “bottom-up” approach. In the latter case it is assumed that purely physical effects determine the dynamics of lower levels of organization and, by extension, strictly govern interactions occurring at higher levels as well. Information, however, can acquire a causally efficacious role in physical systems without violating “the principle of the closure of the physical world” [10]. In fact, an emerging school of thought in evolutionary biology is advancing the hypothesis that the transition from non-life to life, abiogenesis, can be aptly described as a transition in causation and information flow:
“Transitions in causal architecture could have occurred through a series of “information takeover” events, which differ from the notion of “genetic takeover” in that they are not necessarily confined to the transfer of genetic information from one molecular species to another. Instead, information takeover refers to transitions in chemical complexity whereby advances in information processing and computation would have conferred nascent life with the logical architecture necessary to gain control over the chemical substrates in which it was instantiated. These transitions will likely be difficult to identify, however one key feature should be that the (informational) states of a nascent living systems played an increasingly prominent role in determining the future dynamics. This suggests that the processes leading to the emergence of life should be marked by a series of “information take-over” events corresponding to changes in causal architecture and information flow, which, in principle, is measurable.” [9]
Emerging life on Europa, by definition, should have underwent a similar “information takeover” event and this unexplored mechanism by which information is gaining a causally efficacious role in physical systems constitutes the most fundamental convergent evolutionary process imaginable in the realm of biological complexity. Without delving into the philosophical implications of such phenomena too far, I am definitely not viewing “information takeover” events as contingent happenstance. They rather represent necessary transmutations or transitions in the overall complex organization of the observable Universe. I challenge every contingency-oriented naysayer to consider these cutting-edge theories on the origin of life and to provide a coherent theoretical framework in which life can still be explained as a “cosmic accident”.
Informational takeover events appear to have occurred during all of the major transitions in evolution. At any given higher level of biological complexity novel top-down informational hierarchies are defining lower level molecular, cellular or organismal operations. This idea is closely related to the concept of functional equivalence classes:
“Top-down causation operates through functional equivalence classes. Functional equivalence occurs when a given “higher-level” state leads to the same high-level outcome, independent of which “lower-level” state instantiates it. Equivalence classes are defined in terms of their function, not their particular physical instantiation: operations are considered (functionally) equivalent (i.e., in the same equivalence class) if they produce the same outcome for different lower-level mechanisms. Functional equivalence classes therefore represent the physical manifestations of virtual constructors. Functional equivalence is evident, for example, in the case of convergent evolution presented above, where convergence occurs because natural selection optimizes a functionally equivalent outcome (in this case, echolocation).” [9]
Clearly, the proposed MCTs that could potentially constrain the selection of future biosignature detectors constitute a practical application of the top-down causation via functional equivalence class paradigm. Accordingly, each MCT represents a fundamentally organismal functional equivalence class, performing core cellular operations, which can be instantiated, imaginably, by a multitude of different functionally interchangeable biopolymer or metabolic networks. In terrestrial organisms the MCT of molecular replication is instantiated by DNA (although DNA cannot, of course, replicate on its own, it needs RNA and protein “co-factors”). However, for the present purpose it suffices to define DNA as the main molecular information deposit that gets copied during the process of cellular reproduction. Contrary to neo-Darwinian dogma top-down causation can influence genome evolution in a non-random way as evidenced by convergent changes in hundreds of mutations that benefit the functional emergence of a trait that can serve as a direct adaptation to a specific environmental context. The genomic study on echolocation in dolphins and bats [11] referenced in the cited paragraphs above and below is one of a growing number of publications that begin to assess the wonders of convergent evolution on a genomic scale and its impact on the emergence of evolutionary novelties:
“Top-down causation is an important mechanism for adaptive evolution through natural selection, where the higher-level “goal” is survival. The role of top-down causation in adaptive selection is particularly evident for cases of convergent evolution. A striking example is provided by the evolution of echolocation in dolphins and bats, where over 200 genes have independently changed in the same ways to confer both species with the ability to use sonar. A common high-level selection pressure (e.g., for the ability to navigate) lead independently to the same specific mutations in the DNA of both species. Convergent evolution thus provides a clear example of top-down causation via adaptive selection, where causal influences run from macroscopic environmental context to microscopic biochemical structure.” [9]
Another remarkable example how function is shaping genome evolution is the convergent expansion of the proto-cadherin protein family between the invertebrate octopus and vertebrates allowing for increased neuronal complexity in each respective evolutionary lineage, a finding that was revealed by a recent sequencing effort [12]. We are just beginning our quest in understanding how the emergence of a certain biological function, in this case synaptic/neuronal signalling, is “driving” large-scale genomic rearrangements that go well beyond what is commonly understood as “mutational” or “gene duplication” events.
Yet an even more illustrative phenomenon showing how biological complexity is governed by top-down causation is the regulation of gene expression by the so called epigenome. Genomic information is stored in a physical object, DNA. The entirety of DNA in a given organism is constituted by its genome. Nonetheless, the genome does not contain the complete information on how, when and to what extent certain genes should be expressed or repressed. This critical cellular information is instantiated in the epigenome. While the epigenome is associated with certain non-DNA molecular sequences (e.g. histone modifications), it should be rather viewed as a “… global, systemic, entity” whose “… real-time operation …lies in the realm of nonlinear bifurcations, interlocking feedback loops, distributed networks, top-down causation and other concepts familiar from the complex systems theory.” [13].
It should be quite clear to any researcher working in the biosciences today that – quite paradoxically – the “central dogma” of molecular biology (unidirectional information flow from DNA RNA protein) is not compatible with biological reality. Therefore biologists are beginning to think along the lines of “information hierarchies” instead of the “central dogma”. Information hierarchies allow for bidirectional information flow from higher to lower levels of biological organization. Yet, bidirectional information flow does not imply equivalence of each respective structural level in regulating biological systems. For this reason have the causal relationships responsible for the homeostasis of a particular organizational instance been termed “informational hierarchies”, as higher levels are constraining the dynamics of lower levels via information control, processing and selection.
Different molecules become coupled via information selection during biological network constitution as they are carrying out a specific cellular operation. In terms of network topology this process is leading to the formation of hubs, i.e. regulatory molecules forming edges (interactions) with other molecules (nodes) within a group and fewer edges with other groups (that are built around their own particular hub). The result is the hierarchical and modular organization observed in many biomolecular networks, prime examples being gene regulatory and metabolic networks. Hierarchical modular networks display self-similarity: many highly integrated small modules group into a few larger modules, which in turn can be integrated into even larger modules. Consequently, metabolic and gene regulatory networks are grouped into several large modules, which are further partitioned into smaller, but more integrated submodules (for a mathematical description see [14]).
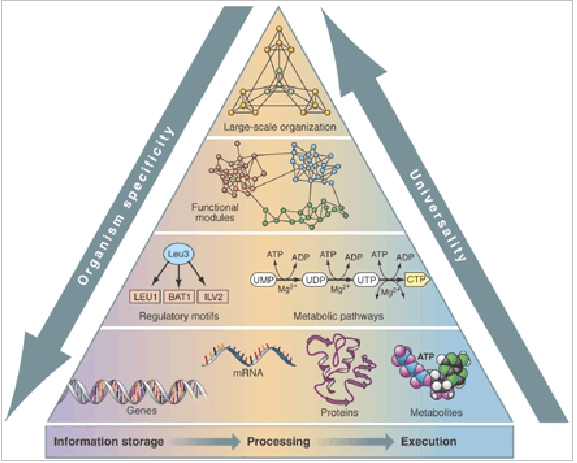
Image: Life’s complexity pyramid. Biological systems are governed by hierarchical, modular biomolecular networks that display large-scale self-similar organization. Organismal molecular information is highly specific while the overall embodied structures encoded within these instructions show recurring patterns universally expressed at any given level of biocomplexity. Figure from (Oltvai and Barabasi 2002) [15]
Lastly, the profound functional similarities in the large-scale organization of biological and technological complex adaptive systems are pointing toward as of yet undiscovered ordering principles underlying physical reality that can be framed within a future integrative framework based on top-down causation and convergent evolution:
“Advanced technologies and biology have extremely different physical implementations, but they are far more alike in systems-level organization than is widely appreciated. Convergent evolution in both domains produces modular architectures that are composed of elaborate hierarchies of protocols and layers of feedback regulation, are driven by demand for robustness to uncertain environments, and use often imprecise components. This complexity may be largely hidden in idealized laboratory settings and in normal operation, becoming conspicuous only when contributing to rare cascading failures. These puzzling and paradoxical features are neither accidental nor artificial, but derive from a deep and necessary interplay between complexity and robustness, modularity, feedback, and fragility.” [16]
ADVANCED BIOCOMPLEXITY ON EUROPA
Next to the water column and seafloor of the deep sea, its underlying sediment presents the largest potential habitat on Earth. On Europa and Enceladus the interaction between the rocky mantle and the overlying water column could lead to hydrothermal processes which may drive the exchange of energy and biogenic elements. In regard to putative hydrothermal systems on these moons, especially in the case of Europa since it is considerably larger than Enceladus, it is conceivable that the previously mentioned serpentinization, a hydro-geological process well-known from certain areas of ultramafic terrestrial seafloor, might provide an energy and chemically rich environment. Thermal cracking of Europa?s crust caused by progressing serpentinization of ultramafic seafloor, a mantle composition that is consistent with models of Europa?s planetary differentiation might reach up to 25 km into the mantle releasing large quantities of molecular hydrogen. This could allow for a putative H2-driven biosphere of microorganisms capable of exploiting chemical rather than solar energy for their biosynthetic and metabolic processes.
Based on these findings we can expand our notion of habitability of icy satellites from the subsurface ocean and seafloor regions down into the mantle itself, where serpentinization reactions potentially fuel a microbial biosphere. Given that this microbial biosphere extends to the seafloor where hydrothermal systems are representing an interface between the ocean and mantle geochemistry, we can start to reasonably assume the existence of simple ecosystems based on chemoautotrophic microorganisms. How this type of unicellular life, often found in a symbiotic relationship with various multicellular species, is giving rise to complex ecosystems has been best studied at hydrothermal vent sites across the mid-ocean ridges of our planet. Global heat output models for Europa have shown that hydrothermal systems could in theory support energy fluxes roughly in the range of terrestrial activity.
Additionally, another important process that might enable the emergence of complex life beyond the unicellular stage on Europa is the transport of radiolytically produced hydrogen peroxide from the surface down into the ocean, where it would disintegrate into H2 and O2. Calculations which linked the amounts of potentially available O2 to the length of the delivery period of H2O2 from the surface to the ocean showed that upper limit abundances of hydrogen peroxide might lead to O2 concentrations above those of terrestrial oceanic O2 minimum zones within a turnover time of 10–110 Ma. Estimates of mean surface age of 30–70 Ma thus allow the possibility of oxygenated upper ocean layers. The latest spectroscopic assessment of H2O2 abundances on Europa?s surface using Earth-based adaptive optic systems lowered high-end estimates but nevertheless allows for considerable amounts of dissolved oxygen inside the ocean if mixing occurs. Further spectroscopic analysis carried out by the same researchers has led to the detection of a previously unknown surface component, the magnesium sulfate salt epsomite. As magnesium is unlikely to be an endogenous constituent of the surface it probably originates from oceanic magnesium-chloride reaching the surface and reacting with previously present sulfate compounds.
A recent paper from the well-known JPL researchers Steve Vance, Kevin Hand and Robert Pappalardo largely confirms the previously calculated estimates of H2 and O2 abundances, thereby stressing again Europa’s potential for the emergence and perpetuation of life in terms of geochemical energy disequilibria over the entire planetary history [17]. Hence it is tempting to consider hydrothermal vents not only as a potential environment for a second genesis of life on Europa, but also to think about them as enduring habitats for biological activity from which simple microbial life is possibly evolving towards higher complexity into multicellular organisms that over time become endogenous to the overlying water column and ice/ocean interface. Convection currents would be able to carry away microorganisms from their native seafloor safe haven into harsher environments where their survival would depend on the evolution of novel energy transduction mechanisms. Given the unprecedented adaptive versatility of microorganisms from Earth and the unforgiving selective pressures within Europa?s dark ocean this does not seem entirely unlikely. Ecosystem modeling approaches for Europa based in one case on methanogenesis, a known pathway employed by terrestrial chemoautotrophs, and in the other on a hypothetical energy transduction mechanism (ionotrophy), has yielded theoretical absolute biomass abundances that would be high enough to sustain complex and differentiated ecological networks comprising producers, primary, secondary and tertiary consumers and detritivores in analogy to terrestrial marine ecosystems [18]. In this model the tertiary consumers and benthic detritivores are comparable in size to terrestrial tadpoles and brine shrimp.
Consequently, the existence of macrofauna on Europa should not be discarded a priori. Quite on the contrary, the emergence of multicellularity, a remarkable example of convergent evolution, within a sustained biosphere of microbial chemoautotrophs seems not impossible at all. However, it should be noted that the emergence of multicellularity, as we know it from Earth, requires another important macroevolutionary step. First of all, cell types with an increased genomic complexity, eukaryote-like organisms, need to arise. Interestingly, on Earth this transition happened only once (thereby being one of the bona fide evolutionary bottlenecks in the emergence of higher-order biocomplexity; however we cannot exclude that now extinct alternative eukaryote-like cell designs might have existed at some point in Earth’s history). Then again, secondary endosymbiotic events among eukaryotes occurred repeatedly in a convergent manner. Subsequently, these complex unicellular organisms, repeatedly and independently, turned multicellular nearly forty times in aerobic environments – a prime example of convergent evolution.
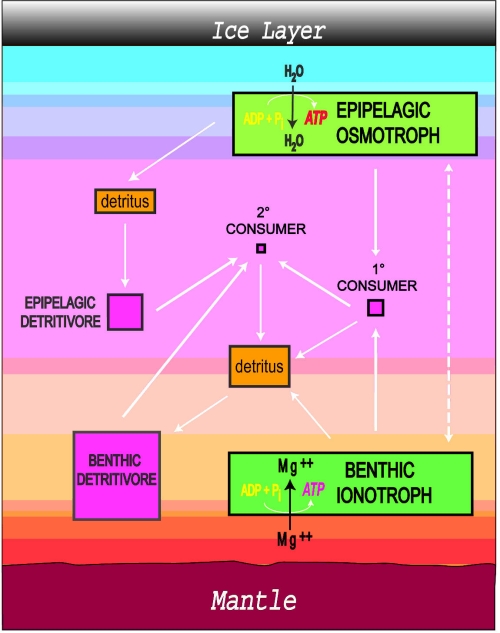
Image: Complex ecosystems might be thriving hidden beneath Europa’s ice shell. Figure from (Irwin and Schulze-Makuch 2003) [18]
LIVING LIGHTS OF THE ABYSS
Contemplating how terrestrial deep-sea ecosystems are organized and which functional adaptations are widely common among marine organisms we are drawn towards the mesmerizing beauty of bioluminescence. As already noted the phenomenon of bioluminescence is one of the most striking examples of convergent evolution. A basic bioluminescent system contains a light-producing substrate, the luciferin, and an enzyme, the luciferase, which catalyzes an oxidative reaction that leads to the emission of photons in the spectrum of visible light with most luminescence between 450 – 635 nm. In the oceanic world most bioluminescence emitted lies in the blue spectrum, since these wavelengths are travelling farthest in water. The particular type of bioluminescent chemical reaction is strictly dependent on the presence of O2. Many groups of bioluminescent organisms share a common substrate for their light-producing biochemical reactions but independently evolved specific enzymes for catalysis. Bioluminescence mediates fundamental ecological functions including the search for food, attraction of sexual partners and evasion of predators. However, not only multicellular organisms are bioluminescent but also unicellular bacteria and eukaryotes.
Ecosystems inside Europa’s ocean might be organized by bioluminescence on a fundamental level. In upper highly oxygenated oceanic layers bioluminescence might be a wide-spread adaptation of multicellular organisms whose survival is dependent on the location of prey and the attraction of species members for reproduction. However, unicellular life on Europa might also be adapted in an analogous way. On Earth bioluminescent bacteria are often found in symbiosis with more complex organisms such as fish and squid. Coevolution has led to highly differentiated organs containing bioluminescent bacteria that can be moved, focused and activated in response to certain stimuli. Symbiosis between uni- and multicellular life would be very likely in a biosphere on Europa since evolution on Earth has been profoundly shaped by such organismic relationships.
The credit for the first ever mention of bioluminescence as a spectral signature of life in astrobiological exploration goes to MIT exoplanet hunter Sara Seager [19]. She suggested that far-future space telescopes might be able to detect the light signature of gigantic bioluminescent blooms on the surface of distant ocean worlds. On Earth vast blooms of unicellular luminescent bacteria are responsible for so called “milky seas” that have been detected via remote sensing from space [20]. In microbial blooms bioluminescence is the product of quorum sensing which represents an elaborated mechanism allowing bacteria to modulate gene expression and eventually population density. However, quorum sensing is coordinated by chemical signaling and the function of the luminescent response in freefloating blooms of microbes remains to be determined. In a fascinating twist one very advanced, but Earth-bound, telescope actually has detected bioluminescence as signature of life already – as a side-product of astronomical observations. The photo-multipliers of the deep-sea neutrino telescope ANTARES, located in the Mediterranean See, recorded seasonal spikes in ambient luminescence that were subsequently attributed to the activity of enormous underwater blooms of a newly characterized strain of deep sea microbes, Photobacterium phosphoreum ANT-2200 [21].

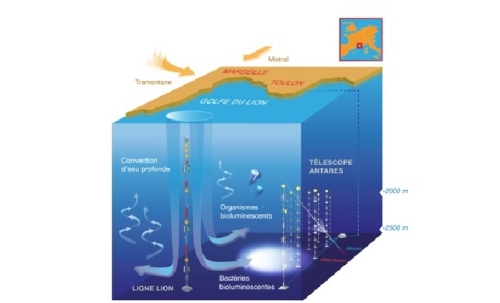
Top image: satellite image of a “milky sea” off the coast of Somalia. Bottom: deep-sea bloom of bioluminescent bacteria detected by the Antares neutrino telescope in the Golfe du Lion, France. Image Credit: Steve Miller/NRL/Mathilde Destelle
Bioluminescence as a new type of biosignature for future exploration of Europa within our own Solar System was proposed by myself at the 64th International Astronautical Congress in Beijing for the first time in 2013. My subsequent theoretical work on this concept was mostly related to assessing the stability – or evolutionary robustness – of bioluminescence as an independently and repeatedly evolved evolutionary trait in phylogenetically distantly related organisms. The latest genomic analyses on early animal evolution confirm that bioluminescence is one of the most ancient functional adaptions in multicellular organisms and a recent study has reassessed the previously underestimated diversity of bioluminescent systems among fishes. As will be explained further below bioluminescence can also serve as an excellent example to explain adaptive evolutionary processes in terms of top-down causation, information hierarchies and functional equivalence classes.

Image: Presenting “Cosmic Convergent Evolution of Bioluminescence on Europa” at the 64th International Astronautical Congress in Beijing, 2013. I am the fifth person from the right, next to former ESA Director General Jean-Jacques Dordain. My conference stay was sponsored by ESA’s “IAC Student Participation Programme”. Image Credit: ESA
The idea of bioluminescence as biosignature is corroborated by novel deep-sea exploration techniques employed in terrestrial oceans that were highly successful within the last decade in identifying a number of previously unknown species. In 2012 the “optical luring” approach developed by Edith Widder allowed for the first detailed video footage of the giant squid, Architeuthis dux, in its natural habitat. This unobtrusive exploration method uses a spherical probe equipped with an array of LED lights that gets lowered into the ocean depths. The probe is called the “electronic jellyfish” because it emulates the spectacular blue bioluminescence pattern of the deep-sea dwelling species Atolla wyvillei. The biomimetic probe is attached to a silent camera system which illuminates the surrounding with far red light because this range of the spectrum is not visible to most deep-sea organisms. Another kind of detection method for bioluminescence has been development in the form of so called bathy-photometers. These devices are made up of flow chambers that are coupled to photomultiplier tubes for measuring the intensity of mechanically stimulated bioluminescence of unicellular organisms. A number of modularized off-the-shelve bathy-photometers is available and can be attached to the latest AUV platforms. This allows for long-range, uninterrupted bioluminescence measurements in varying depths. Another platform that has been developed with the dedicated goal of exploring bioluminescence in deep waters (up to 300 m) is the “Exosuit”, a kind of wearable submarine. Bioluminescence systems of undiscovered species might be used in future medical diagnostic procedures. It made its maiden voyage during an underwater archaeology expedition to the Antikythera site, Greece, in 2014.

Image: The “Exosuit”: taking the spacewalk experience underwater. Image credit: Brett Seymour
On Europa, a submersible robotic exploration vessel could be equipped with highly miniaturized bathy-photometers that would scan for ambient bioluminescence primarily emitted by microorganisms while the optical luring approach, based on its terrestrial counterpart, would target complex multicellular life. Biologically produced light is a far more readily detectable signature of life than for example complex biopolymers are. The biological origin of these compounds can only be verified through multistep chemical analyses, which always bear the danger of a false positive. Thus, the presence of living light inside the ocean of Europa is the most unambiguous sign of biological activity. It can emerge in both uni- and even multicellular organisms as a necessary outcome of the evolutionary process that is determined by the planetary properties of Europa (submarine alkaline hydrothermal vent origin of life and ongoing oxygenation throughout its history).
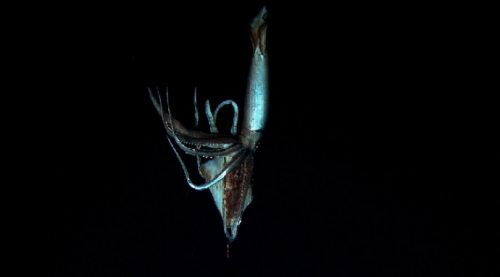

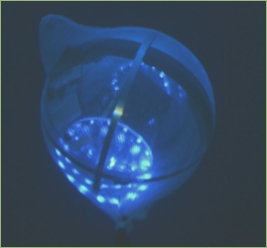
Top image: still from the video that captured the giant squid for the first time. Middle: successful luring of the deep-sea creature with the biomimetic probe. Bottom: “electronic jellyfish” emulating bioluminescence patterns. Image Credit: Reuters/Discovery Channel/Edith Widder
Until now, however, the evolutionary origins of bioluminescence in terrestrial oceans are not completely resolved. One well-founded hypothesis, initially posited in the 1960s, suggests that biological light had its beginning as an ancient oxygen defense mechanism following the onset of the first “great oxidation event” (GOE) approximately 2400 Ma ago. Before the continuous rise of atmospheric and oceanic O2 concentrations due to the evolution of oxygenic photosynthesis in ancestral cyanobacteria, all life on Earth had an anaerobic metabolism. Concentrations of O2 remained relatively low for about 1600 Ma until 800 – 400 Ma ago levels reached half or more of present-day values. This drastic increase in available oxygen occurred simultaneously with the Cambrian Explosion during which the greatest diversification of terrestrial life so far took place. When the ancient oceans were oxygenated in small amounts for the first time after the GOE, chemically reactive O2 species like hydrogen peroxide, singlet oxygen, superoxide and oxygen radicals formed which were highly toxic to the then extant life. It is possible that functional luciferin/luciferase systems evolved during this time, consuming ambient excess oxygen, to prevent the adverse effects of the newly introduced oxidative agents. As a nonfunctional by-product of these reactions bioluminescent light was produced. Within the ancient oceans of Earth all organisms would have been exposed to the same selective pressure, the threat of damaging oxidative reactions, and different species probably employed unrelated chemical systems in their individually evolving bioluminescence.
Once again, we can turn to the previously elaborated theoretical framework underlying convergent evolution via top-down causation. In the same way that MCTs should encompass fundamental systemic properties of alien life forms that could potentially be instantiated by alternative biochemistries, bioluminescence represents an envisioned stable functional state beyond the “minimal convergent” level of biological complexity. Specifically, it is dependent on the presence of oxygen inside the ocean of Europa. If our models on Europa’s geochemistry hold true, ongoing oxygenation would have surely presented a high-level selective pressure in the form of rising oxygen concentrations. This would allow for an evolutionary scenario “where causal influences run from macroscopic environmental context to microscopic biochemical structure” [9]. Thus, such causal influences would lead to the convergent evolution of bioluminescence across Europan biota through top-down causation via adaptive selection. The production of biological light would present a higher-order functional equivalence class that could be instantiated by a number of different biochemical reaction systems, as evidenced by the disparate mechanisms employed by terrestrial organisms. On one hand, potentially further constraining the exact biochemistry underlying bioluminescence on Europa could be a future research goal; on the other, future biosignature detectors for biological light can be designed irrespective of the lower level operational molecular embodiment. Therefore, in a very practical but also in an almost mystical sense, bio-photonic light appears to transcend its particular substratum.

Image: Phylogenetic distribution of marine bioluminescence across a number of uni- and multicellular phyla. This scheme nicely illustrates how bioluminescence as an adaptive organismal trait is linked to the concept of functional equivalence classes. Biological light can be instantiated by a number of different chemical systems based on, sometimes, shared substrates but individually evolved catalytic enzymes: the essence of convergent evolution. Figure from (Widder 2010) [22]
After the first one and a half billion years or so after the GOE, probably only bacterial microorganisms were adapted in such a way since eukaryotes had not yet entered the evolutionary scenery. Still, luciferases could have provided an adequate oxygen protection mechanism for these more complex cells as well. After all, the specific luciferases of modern-day organisms like jellyfish, for example, are not related to bacterial enzymes. Accordingly, their bioluminescent systems emerged independently from each other by way of convergent evolution. Sometime before the Cambrian Explosion, after the oceanic oxygenation had immensely progressed and incipient multicellular animal life did not rely on luciferases as their primary oxygen defense anymore (more efficient means of detoxification certainly evolved in parallel), a decisive shift of selective pressure secured the fate of bioluminescence as a functional adaptation. Luciferin/luciferase systems would have lost their selective value and eventually been lost in the presence of equally or more effective oxygen defense if not another function of the biologically produced light was subsequently being selected for. Obviously, such co-opting of the function of bioluminescence depended on the pre-existence of light-detecting or rudimentary visual systems capable of detecting the emitted light. How could such biological adaptations, which on Earth evolved in constant interaction with the solar flux, emerge in the pitch-black abyss of Europa?
Light-detecting sensory adaptations are very common throughout the animal kingdom and the underlying mechanism is highly conserved – from the eyespot of the algae Euglena to complex imaging camera eyes of humans and octopus. Vision is always mediated by photoreceptor cells employing rhodopsin-like photopigments linked to G-protein cascades. Photon-induced cis-trans isomerization of retinal (opsin + retinal = rhodopsin) is the first step in an elaborated second messenger pathway which is responsible for the transduction of the sensory stimulus into electrical signals. Functional photoreceptors using this kind of light-detection and phototransduction apparatus were already in place more than 600 Ma ago before the major branching of animal phylogeny. The basic mechanism underlying light-detection, however, has been repurposed independently several times in the evolution of animals. This lead to the emergence of eyes in jellyfish (in rudimentary form) and complex camera eyes in invertebrates (e.g. octopus) and vertebrates. Hence, on Earth, the function of bioluminescence could be slowly transformed from oxygen defense to its modern-day role in cross- and interspecific communication, predatory evasion and propagation. Although the explanation of the evolutionary origins of bioluminescence as an ancient oxygen defense does not seem to be valid for all forms of ecologically functional light, it accounts for a number of important features observed in extant luminous species: every bioluminescence without exception is depended on oxygen; the global influence of the GOE presented a universal selective pressure to all extant life and thereby led to convergent evolution; bioluminescence is predominantly found among marine organisms since luciferases started to turn into beneficial adaptations at a point in time when all life was still oceanic.
Recent genomic analysis of the comb jelly Mnemiopsis leidyi and Pleurobrachia bachei suggests the phylum of ctenophores, or comb jellies, (of which most members are bioluminescent) as the most basal group of metazoans [23]. Certain features in the genome structure of Mnemiopis are even implicating the co-evolution of emerging multicellular organization and the origin of coupled light-producing and detecting proteins. Thus, early vision systems could have possibly emerged in the deep sea for the first time or at least independently from pelagic organisms and their sunlit habitats. In fact, the newly released comb jelly genomes have caused a major controversy among evolutionary biologists because their primary branching off from the animal tree of life would either indicate a far more complex last common ancestor of all animals or convergent evolution of the most complex adaptation in nature: nervous systems. Both evolutionary scenarios pose serious problems to the neo-Darwinian doctrine. My current research is dealing with this fascinating evolutionary conundrum and I am currently working on comparative genomic analyses of the neurotransmitter complement of each respective animal phylum to discern whether neurons evolved only once or multiple times. In any case, yet another putative route to vision in sunless environments is presented by “infrared” vision as observed in the deep-sea shrimp Rimicaris exoculata. This species has evolved specialized cellular structures to detect the thermal signatures of hydrothermal vents. I hope to elaborate on these issues in a forthcoming paper that will specifically deal with the convergent emergence of bioluminescence on Europa. For now it suffices to clearly state that there are known adaptive routes among terrestrial biota enabling them to evolve visual systems in the absence of light. Consequently, on Europa, there might exist molecular pathways to not only produce bioluminescence but also to detect it. Ensuing co-evolution of both functions would obviously lead to a kind of amplifying “run-away” evolutionary process related to bioluminescent systems complexification. Amazingly, inside Europa’s abyssal ocean, light could have potentially been turned into a biological resource determining ecosystem dynamics beyond the organismal scale.
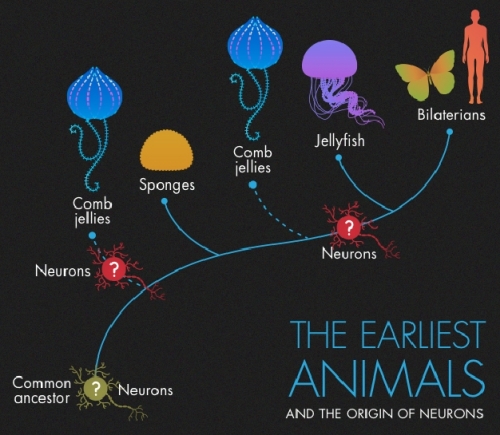
Image: The first sequenced comb jelly genomes are revolutionizing our understanding of the evolutionary emergence of animal complexity. Even complex nervous systems could be a product of convergent evolution. Image credit: Olena Shmahalo/Quanta magazine
What, then, can we learn from the evolutionary history of bioluminescence on Earth in regard to its hypothesized convergent emergence on Europa? First, oxygen is apparently indispensable for the evolution and maintenance of bioluminescence. In the ocean of Europa repeated oxygenation events could potentially occur due to the transport of oxidants from the surface. Depending on the exact mechanism of surface/ocean communication either a continuous and slow oxygenation or a more dramatic and periodically occurring influx of oxygen seem plausible. In any case, putative organisms on the rather hypoxic or anoxic seafloor that are pioneering in the exploration of the more oxygenated upper ocean layers would be in need of efficient means of oxygen detoxification. The convergent evolution of bioluminescence on Earth is a proof of the evolutionary robustness of luciferin/luciferase systems in achieving such adaptations in a global oceanic habitat. Secondly, rudimentary visual systems must have had already evolved (in multicellular organisms) as a pre-adaptation before the coopting of non-functional biological light, resulting from the activity of luciferin/luciferase systems acting as a means of oxidative protection, into ecologically relevant bioluminescence could begin.
CONCLUSIONS
The main idea behind this essay was to promote a new representation of evolutionary biology, one that is more encompassing than its predecessor – the neo-Darwinian synthesis. Based on the theoretical and practical work of many colleagues I termed this paradigm “Cosmic Convergent Evolution”. Like any scientific theory, CCE is a model, an attempt to understand the complexities of physical reality within the limitations of the human mind. Previous and current assumptions, hypotheses and mechanisms dealing with the origin, evolution and development of complex biological systems can be reassessed through the lens of CCE – evolutionary science always had an inherently retrograde character to it. However, in the grand vision of bioluminescent life forms on Europa I see hidden the predictive powers of CCE. If the combined body of scientific theorizing, experimental work and data streams acquired from decades of robotic space exploration hold true we have good reason to believe that a light is shining in the abyssal depths of Europa.
Fascinatingly, in a kind of self-referential meta-convergence, CCE does not only offer novel explanatory avenues for the emergence of biocomplexity. Technological applications, too, can be derived using its underlying assumption that top-down causation, convergent evolution and modularity are principles associated with biological as well as technological complex adaptive systems. Astrobiology is going to be the scientific and technological testing ground for CCE. The blue spectres of abyssal Europa are awaiting their discovery. Prometheus will be unbound.
Validation or falsification of CCE’s main tenets can come from disparate areas of research, be it the origin of life field, artificial intelligence, future space exploration, SETI studies, or animal cognition. Having experienced the wonders of convergent evolution myself by encountering the mesmerizing sentience of spotted and bottlenose dolphins in the Bahamas last year, my personal bet is on a dedicated organismal perspective. Research like that of Dr. Denise Herzing of the “Wild Dolphin Project” who is trying to crack the code of dolphin communication is highly successful in furthering our understanding and appreciation of “alien” intelligences. Vast amounts of information is still hidden in the genomes of countless unexplored species that inhabit our beautiful terrestrial ecosystems, many of them in our oceans. Future sequencing efforts of animal complexity and also metagenomic studies of microscopic life forms will revolutionize our understanding of genome evolution and the evolutionary process as a whole.
To summarize: Explaining the origin of life, pre-cellular evolution and multicellular biocomplexity in purely (neo)-Darwinian terms is like describing the quantum world with Newtonian physics. It is easy to see that “Universal Newtonism” is not able to unify modern-day physics; why should “Universal Darwinism” be able to do so when we consider the ever-increasing non-linear hyperspace of biological complexity [24]?
Personally, I never fully understood how Darwinism is purported to “integrate” all of the existing biological sciences. Among biologists it is widely acknowledged that The Origin of Species didn’t even satisfactorily explain its main subject, namely the process of speciation. As science progresses, we have to let go of much cherished models (i.e. the neo-Darwinian Synthesis) that do not suffice anymore to explain the phenomena we are theorizing and observing. Or to use a biological analogy: a kind of endosymbiosis takes place, where an advanced cell takes up a more limited-complexity unit, and eventually integrates it into its own cellular context where it will serve an important but subordinate function (given it is still viable within the new systemic whole; degeneration and disintegration is always an option too). This is simply how science works. Hard-core Darwinists (again, why are there no “Newtonists” around anymore?) shouldn’t fear some evil machinations of the creationists they so much love to vilify. They should rather watch out for the impact of overwhelming amounts of new biological data, as well as novel theoretical concepts, on their cherished notions of “Darwin-fits-it-all” evolution. In my opinion, orthodox understandings of evolutionary theory simply fail in explaining the observation of emerging higher-order systemic complexity throughout every major evolutionary transition. Any other approach apart from letting go of previous world-views will become dogma and ultimately earn its deserved spot in the dustbin of history. Again, Let us turn to the eternally echoing words of the eminent Carl R. Woese:
“Science is impelled by two main factors, technological advance and a guiding vision (overview). A properly balanced relationship between the two is key to the successful development of a science: without the proper technological advances the road ahead is blocked. Without a guiding vision there is no road ahead; …” [1]
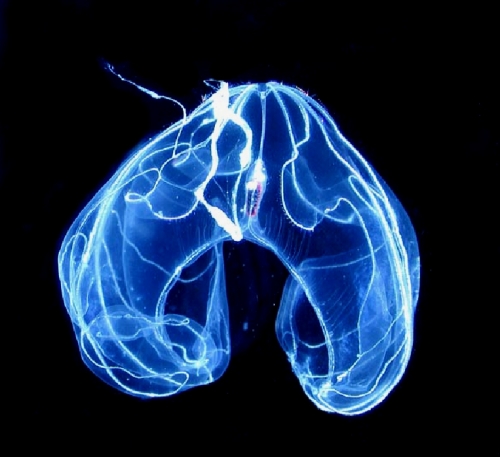
Image: Terrestrial comb jelly: Bathocyroe fosteri. Credit: Wikimedia Commons.
The Europan specimen was contained in a sealed-off translucent tank that was linked to a fresh reservoir of previously harvested icy moon ocean water, which was to be constantly pumped into the micro-habitat. Failure in the form of a dead alien creature delivered to Earth was not an option. Castor had been sitting here in the ship’s bio-hazard compartment for hours – or days? The pulsating patterns of light seemingly synchronized with his own waves of neural activity, pondering the depths of the alien consciousness in front of him. It was aware, it was moving, it was reacting, it was thinking – that was a certainty for the astrobiology officer. Unfortunately, any preliminary investigations, molecular or behavioral, had been strictly prohibited by Military Intelligence. He was lucky enough to have been able to sneak into this area and at least spend the time of the voyage with his discovery. He had brought his personal sim device with him on which he had compiled all the available bioluminescent patters recorded from Earth’s oceans. This data was collected by the underwater drones that had been autonomously exploring the terrestrial abyssal regions for years, preceding the Solar System expedition. “Let’s see what this deep learning baby can do for me” he thought and held his device in front of the tank to capture the alien light emanations. “Finally I can use this camera to capture something even more stunning than the Miami beach vixens from last year’s summer holiday” he laughingly joked to himself standing up in front of the calmly floating jelly that was producing a wonderful diaphanous color display. In reality he had spent his last three summers just above the bottom of the Pacific, eleven kilometers below the surface, at Mariana-5 underwater station to exercise the upcoming EVAs. A time spent well in the light of his successful operations on Europa.
“Hey A(I)migo, please compare these light patterns emitted by our little fellow here to the deep sea bioluminescent flash database. Yeah, you know, the one that I compiled from all the AUV detectors.” A moment of silence. “Access denied. No security clearance granted for this dataset.” In desperation Castor thought that even he wasn’t able to hack into the quantum-encrypted Global Military Intelligence Defense Network. After all, it was designed to withstand potential cyberattacks carried out by alien intelligences. Suddenly, rhythmic bright flashes started to throb through the extraterrestrial jelly. It swam closer to the wall separating Castor from the 4° C cooled analogue environment on the other side. While it was spreading its gelatinous wings in a dramatic movement of embodied sentience the electronics went haywire.
The lights in the gigantic hall went out and everything was enshrouded in the gleaming epiphany of spectral blue luminescence. Castor’s handheld A(I)migo went online again. “Access granted. Search is commencing, Sir”. A moment of silence, again, only interrupted by the reassuring beeps highlighting the successful matches of the neuronal-augmented pattern recognition algorithm working with the speed of light. Projections began to appear on his contacts, geometric shapes representing the principal component analysis of the informational content encoded in the bioluminescence – – convergent forms, most beautiful, from Earth and Europa. Castor smiled: “Okay, let’s see what we have here.”
Acknowledgements: I am greatly indebted to my twin brother, Danilo, for inspirational discussions and ever supportive editorial help. My gratitude also goes to Maziar and Jahangir Demahsi for helping in the production of our no-budget documentary film “KONVERGENZ”!
Author contact: claudio.flores.martinez@embl.de
ENDNOTES
1. Woese, C.R., “A new biology for a new century.” Microbiol Mol Biol Rev, 2004. 68(2): p. 173-86.
2. Flores Martinez, C.L., “SETI in the light of cosmic convergent evolution.” Acta Astronautica, 2014. 104(1): p. 341-349.
3. Walker, S.I., H. Kim, and P.C. Davies, “The informational architecture of the cell.” Philos Trans A Math Phys Eng Sci, 2016. 374(2063).
4. Smart, J.M., “The transcension hypothesis: Sufficiently advanced civilizations invariably leave our universe, and implications for METI and SETI.” Acta Astronautica, 2012. 78(0): p. 55-68.
5. Russell, M.J., et al., “The drive to life on wet and icy worlds.” Astrobiology, 2014. 14(4): p. 308-43.
6. Flores Martinez, C.L., “Convergent evolution and the search for biosignatures within the solar system and beyond.” Acta Astronautica, 2015. 116: p. 394-402.
7. Chela-Flores, J., “Testing evolutionary convergence on Europa.” International Journal of Astrobiology, 2003. 2(04): p. 307-312.
8. Konstantinidis, K., et al., “A lander mission to probe subglacial water on Saturn?s moon Enceladus for life.” Acta Astronautica, 2015. 106(0): p. 63-89.
9. Walker, I.S., “Top-Down Causation and the Rise of Information in the Emergence of Life.” Information, 2014. 5(3).
10. Auletta, G., G.F. Ellis, and L. Jaeger, “Top-down causation by information control: from a philosophical problem to a scientific research programme.” J R Soc Interface, 2008. 5(27): p. 1159-72.
11. Parker, J., et al., “Genome-wide signatures of convergent evolution in echolocating mammals.” Nature, 2013. 502(7470): p. 228-31.
12. Albertin, C.B., et al., “The octopus genome and the evolution of cephalopod neural and morphological novelties.” Nature, 2015. 524(7564): p. 220-4.
13. Davies, P.C., “The epigenome and top-down causation.” J R Soc Interface, 2012. 2(1): p. 42-8.
14. Ravasz, E., et al., “Hierarchical organization of modularity in metabolic networks.” Science, 2002. 297(5586): p. 1551-5.
15. Oltvai, Z.N. and A.L. Barabasi, “Systems biology. Life’s complexity pyramid.” Science, 2002. 298(5594): p. 763-4.
16. Csete, M.E. and J.C. Doyle, “Reverse engineering of biological complexity.” Science, 2002. 295(5560): p. 1664-9.
17. Vance, S.D., K.P. Hand, and R.T. Pappalardo, “Geophysical controls of chemical disequilibria in Europa. Geophysical Research Letters, 2016. 43(10): p. 4871-4879.
18. Irwin, L.N. and D. Schulze-Makuch, Strategy for modeling putative multilevel ecosystems on Europa.” Astrobiology, 2003. 3(4): p. 813-21.
19. Seager, S., M. Schrenk, and W. Bains, “An astrophysical view of Earth-based metabolic biosignature gases.” Astrobiology, 2012. 12(1): p. 61-82.
20. Miller, S.D., et al., “Detection of a bioluminescent milky sea from space.” Proceedings of the National Academy of Sciences of the United States of America, 2005. 102(40): p. 14181-14184.
21. Tamburini, C., et al., “Deep-Sea Bioluminescence Blooms after Dense Water Formation at the Ocean Surface.” PLoS ONE, 2013. 8(7): p. E67523.
22. Widder, E.A., “Bioluminescence in the ocean: origins of biological, chemical, and ecological diversity.” Science, 2010. 328(5979): p. 704-8.
23. Moroz, L.L., et al., “The ctenophore genome and the evolutionary origins of neural systems.” Nature, 2014. 510(7503): p. 109-114.
24. Morris, S.C., “The navigation of biological hyperspace.” Int J Astrobiol, 2003. 2(02): p. 149-152.



A fascinating, tour de force read. Glad to know that someone is really thinking ahead before we literally plunge into that alien ocean circling Jupiter. The idea of using lights to attract potential native sea creatures – simple yet brilliant.
Of course the native Europans may end up responding to different stimuli, such as smell or sound (Earth oceans are very noisy places, and I am not talking about human ships and boats here), but a series of multicolored lights operating in different patterns and wavelengths should not be too difficult to have as one of the payloads.
Just this morning I came across by chance this article I link to next. In addition to liking its overall message very much, it can also apply to how we approach studying and thinking about potential Europan life forms:
http://www.pnas.org/content/112/14/4185.full
Here is a NASA concept of a swimming robot explorer for Europa from 2015:
http://www.discovery.com/dscovrd/space/will-nasa-send-a-swimming-robot-to-explore-the-oceans-of-europa/
To quote:
“…engineers have proposed a soft-robotic rover, capable of exploring foreign environments, such as the depths of subterranean ocean housed on Jupiter’s moon, Europa. The bot derives its power from the local magnetic fields found on other celestial bodies.”
Here are details on several instruments designed to detect life from a probe that will flyby Europa in the 2020s:
http://www.swri.org/3pubs/ttoday/fall15/eyesoneuropa.htm
Let us just not make the mistake of abandoning Europa in the event that whatever probe we do send into that distant ocean does not detect signs of life, or should I say the kinds of life we were expecting. Remember what happened with Viking when it did not automatically detect the Martian microbes everyone was expecting, including and especially the mission scientists. Europa is almost the size of Luna, has twice the amount of liquid water as Earth does, and the ocean bottom is 60 miles deep (Earth’s deepest trench is only 7 miles down). One probe mission is not going to answer all our questions.
And imagine this kind of encounter at Europa:
https://www.youtube.com/watch?v=SkBpummjR5I
Why the human race isn’t sending multiple missions to Europa and all the other fascinating places in the Sol system (and beyond) shows our cultural parochialism and the limits of our public education system. It is not due to either a lack of funds or technical knowledge. If we could spend one trillion dollars a month on an ultimately futile war, we can afford a few space probes.
I published a speculation along these lines a few years ago in the Futures column of Nature:
http://www.nature.com/nature/journal/v499/n7456/full/499120a.html
…but it is fascinating to see this possibility explored in so much more depth here, and with such richly imagined scientific detail. Great article!
Europa is such a fascinating world. I sure hope we get to take a peek under that ice within my lifetime…
Seeing as we were supposed to get to Mars by 1985, I doubt that. Just before the moon landings the authorities said Mars was next and men would land there by 1985. I was 14 then, now I’m 62 and no Mars landing
in sight yet.
SpaceX may put humans on Mars in 2025, at least one decade earlier than NASA’s current vague goal. Elon Musk hopes to launch an unmanned version of Red Dragon to land on the Red Planet as early as 2018.
http://www.spaceflightinsider.com/organizations/space-exploration-technologies/cost-details-revealed-spacexs-red-dragon-mission/
Hi Ken,
I am tracing the “deep homology” of the bioluminescence idea back to James Cameron’s 1989 “The Abyss”. Assuming most of the readers have seen this movie I am just reminding you all of the scence in which Virgil Brigman (played by Ed Harris) is taking the plunge into the abyss, half-dead, and has an encounter of the third kind, so to speak.
I came up with my bathy-naut scenario already in late 2012 using it for my ESA scholarship application and IAC abstract in the spring of 2013. I presented my paper in September 2013. Having found your brilliant piece some time in 2014 I am lead to count this as a “multiple” in regard to the relative simultaneous appearance of ideas that are “out there”.
Another instantiation of the bioluminescence idea is presented by the creature in “Europa Report” (this specimen being a brain child from Kevin P. Hand and Steve Vance of JPL).
Seriously, convergent evolution does not only apply to the realm of technology but also to the realm of ideas (think Darwin and Wallace as the prime example)
Best regards,
Claudio
Hi Claudio. Congratulation on your work. Would you be able to post a link to your paper here for all of us to benefit from? Thank you.
Claudio,
Director James Cameron obviously loves the idea of bioluminescent creatures, if only for entrancing movie effects. Avatar had a number of such creatures as part of the Pandoran biosphere.
As always, Arthur C Clarke had a fascination too with such glowing creatures. From his early stories of squid communicating with bioluminescence, to the bioluminescent hints of life in an ice cave on Halley’s Comet in “2061: Odyssey 3”.
In real life, there has been some excellent video of such animals from the ocean abysses taken by ROVs. There has also been renewed interest in different colored fluorescent proteins for imaging. We’ve gone from the old staple of GFP from jellyfish to a rainbow of colors sourced from different organisms. There was recently even work on using such proteins in low energy lasers.
You and others may find this article (and the novel) of high interest and importance when it comes to seeking out aquatic alien life:
http://www.pnas.org/content/112/14/4185.full
There has been criticism that the NIH in particular doesn’t fund enough original research, but rather safe research that will likely generate the results expected. We also need public science funding not be be viewed as a financial ROI as that leads to a VC mentality – getting early payback for the capital invested. However we also need some constraints on research that is funded to prevent frittering away funds on ideas that are extremely fringe in the hope that a few might hit pay dirt. There has to be a balance.
As regards Nasa. I find it dispiriting that it has become a pork barrel works program for Congressional districts. The SLS is a good example (as Ares 1 and 5 preceding it) of a waste of money that could be better spent on different projects. It’s high time that manned spaceflight hardware was built by innovative private companies and that Nasa focused on developing new technologies, path finding missions and science that includes the “search for life” as one piece of that science mission.
Because if NASA doesn’t do all those things, someone else will.
http://gbtimes.com/china/china-unveils-its-mars-2020-probe-and-science-goals
A very good story, but do you think it will be humans doing such exploration in person or an Artilect by the time such missions are allowed to happen at Europa and elsewhere in space?
I know it has been felt that such space adventures need the human element, but as we progress with artificial intelligence and robotics, humans may eventually only be required in space for colonization purposes. Look at how the public anthropomorphizes our current and past robotic space explorers which are nowhere near the level of intelligence and sophistication that we will see in the coming decades with real Artilects. Three big examples are known as Spirit, Opportunity, and Curiosity:
https://xkcd.com/695/
When Wernher von Braun envisioned manned lunar missions in the early 1950s, he had huge ships with 75 men for the expedition. He was thinking of and influenced by earlier naval voyages of discovery, which normally required dozens if not hundreds of men to perform all the necessary tasks. He did not anticipate that computers and other machines would be able to do most of the work at the Moon. They would save vast amounts of money, time, and resources that would have been eaten up by having to provide for the needs of dozens of organic crew members.
http://www.thespacereview.com/article/968/1
https://www.youtube.com/watch?v=1ZImSTxbglI
If the goals of Project Apollo had truly and foremost been about the peaceful, cooperative, and scientific exploration of the Moon, we could have and would have done it with machines alone. Five Lunar Orbiters had imaged 99% of the lunar surface in just two years. Five Surveyors (out of 7) had successfully landed on the Moon and each returned thousands of images and data about our celestial neighbor for weeks (Surveyor was also envisioned being equipped with a small rover, but that did not come to pass).
Note how often Surveyor was downplayed in the various histories of direct lunar exploration, because otherwise the question of just how much a human was required to be there to do the same things would be raised. Add in the fact that the Soviet Union did return regolith samples with their later automated Luna landers and explored the Moon with two robot Lunokhod rovers for months and it becomes obvious that the automated study of the Moon could and was done – and far less expensively and with no danger to human life.
http://www.universetoday.com/60142/did-you-know-russian-lunokhod-rovers-created-memorials-on-the-moon-honoring-women/
I know you can say that the Apollo astronauts returned far more lunar surface samples than all of the Soviet Luna missions together by far, but in the end it is quality over quantity. And a more concentrated and cooperative automated lunar program using the machines described above could have done just as well and more, plus they did and could have functioned far longer than trying to keep even a small group of humans alive on the Moon.
As I said above, as AI and robots become even more sophisticated and can conduct in situ analyses of the world they are exploring, the need for humans in the mix will only make sense if it is part of a colonization effort. Especially when we head out into the wider Milky Way galaxy.
Yes, in the best case scenario, animals could arise on Europa closer to the
bottom of the ocean. But life that complex is highly dependent on high powered chemical reactions that yield abundant energy for such animals.
On earth, when mitochondria “invented” the ATP molecule It was a
Revolutionary, far more so than the Cambrian Explosion. In the ATP reaction one of the by products is CO2. If there were ATP like processes on Europa (assuming is it able create free o2, as part of the process), if there is strong upwelling from deep hotter crust to near the ice cap, we should see
Some CO2, continually ‘fizzing’ out of the ice trenches of Europa, while it is close to the liquid layer source. Unfortunately Europa is still a very cold place and if the CO2 strays only a short distance from it’s point of origin It will likely freeze in the very near vicinity or in the ice trenches on the surface.
Now currently abiotic (most likely) oxygen is the only atmospheric component measured at 1 x 10^-12 concentration compared to Earth.. We therefore cannot be sure if there is some Frozen CO2 amounts on the surface. With a lander that can travel near/in the ice trenches, we could seek answers, a mass spectrometer in close orbit MIGHT be able to detect CO2 snow. But the current JUNO mission is not tasked with doing any Europa Science that I know of.
Even if there is no CO2, near the ice trenches, finding any chemicals other than 02, H2, H20, will be a strong hint that Something interesting is going on deep in the Europa’s Hydrosphere. Hopefully NASA is will take up the
challenge of a full investigation of Europa.
Congress told NASA to get their act together about exploring Europa back in May, so maybe:
http://arstechnica.com/science/2016/05/house-budget-provides-260-million-for-two-life-tracking-europa-missions/
and…
http://kutv.com/news/nation-world/life-on-jupiters-moon-europa-congressman-thinks-nasas-mission-can-prove-it
and…
http://www.npr.org/sections/thetwo-way/2016/07/05/484263172/what-comes-next-for-nasa-after-juno-not-much
and…
http://www.aerospaceamerica.org/Documents/Aerospace_America_PDFs_2016/June2016/Feature1_AmbitionEuropa_AA_JUN2016.pdf
and…
http://www.thespacereview.com/article/3025/1
And I highly recommend this blog called Future Planetary Exploration, though I am a bit concerned it has not been updated since early June:
http://futureplanets.blogspot.com/
I am fascinated by the idea of bioluminescence on Europa, but if we find it I will not regard it as evidence of a grand cosmic tendency. Convergence is just a way of saying that there are some obvious good ideas which turn up time and again in evolution, and that does not stop it being contingent.
This is the flaw in the use of convergence as a rebuttal of Jay Gould’s thesis that the emergence of intelligence on earth is purely contingent. If it would have happened anyway because something else would have converged on intelligence anyway, why do we see no actual sign of that convergence happening anyway? Nothing is obviously preventing it. We know that some things are more converged on than others, like sight and bioluminescence, and there is no a priori reason to promote convergence to a universal principle, rather than something that sometimes, purely contingently, happens.
I would add that SETI by looking for radio waves depends strongly on putting an awful lot of faith in convergence. Not saying we shouldn’t do it, as long as it is the only game in town, but “all intelligent life produces radio waves” is no more obviously true then “all flying creatures use echolocation”.
Hi Robert,
Stephen J. Gould’s work is steeped in the overcome notions of the modern synthesis. Nothing exciting to discover there in my opinion (and of those who care to advance evolutionary science to find explanations for the very complexity we are observing).
This is another excellent essay by Woese and Goldenfeld detailing the miserable state of evolutionary biology in the 20th century:
“This is the story of how biology of the 20th century neglected and otherwise mishandled the study of what is arguably the most important problem in all of science: the nature of the evolutionary process. This problem has suffered the indignity of being dismissed as unimportant to a basic understanding of biology by molecular biology; it went effectively unrecognized by a microbiology still in the throes of trying to find itself; and it became the private domain of a quasi-scientific movement, who secreted it away in a morass of petty scholasticism, effectively disguising the fact that their primary concern with it was ideological, not scientific. Despite this discouraging beginning, our story will end well: the study of the microbial world at the beginning of the 21st century is liberating biology from the Procrustean bed of dogma on which it has been cast for so long, and a new understanding of evolution as a process is already beginning to form, in a manner that will eventually supersede the scientifically stultifying language-culture of the 20th century.”
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2650883/
Contingency oriented thinkers are adhering to this doctrine because they are afraid of a creationist takeover of science or something like that. As I am pointing out in my essay, they should be rather concerned with what is happening in actual biological investigation. People like Gould and Dawkins are not contributing to science but rather to the propagation of a certain quasi-scientific, faith-based ideology (yep, exactly the kind of mental attitude they are always criticizing) that seeks to supplant any scientific enquiry that is not based on late 19th century reductive materialism. Maybe it’s because I am coming from an organicist German natural philosophy background, but I am finding these never-ending “skeptic” tirades are so counter-productive in our unbiased “Anschauung” of the natural world. Okay, you are free-thinking, somewhat witty atheists… Got it. Doesn’t immediately turn you into an excellent evolutionary biologist.
I would like to hear how you explain two independent origins of life and accompanying “informational” takeover events as contingent happenstance without introducing “universal principles”. Convergent evolution on the genome level is proving basic tenets of Darwinism wrong, in so far that mutations are not random. Organisms are plastic and their celluar metabolic structure is able to adapt in real time and feedback these changes to the genome. These are topics at the frontier of science but in 20 years people will look back and view evolution according to the neo-Darw. synthesis as a special case of a much vaster, more complex evolutionary process.
The emergence of intelligence is also obviously not “contingent” as neurons have evolved multiple times in animals. It is a necessary condition for the complexification of cognizent beings. In turn, cognition and memory in biology can also be found in non-neuronalized systems, e.g. amoebas and autocatalytic metabolic networks.
I agree to your position on SETI, but technological convergence can also be viewed in more general terms, for example in the cited work of John Smart from my essay.
Life is embodied information and as of now we don’t know how information can acquire causal efficacy in a material substrate. Saying it just happened by accidence is a worldview and not a scientific hypothesis
Best regards,
Claudio
“Convergent evolution on the genome level is proving basic tenets of Darwinism wrong, in so far that mutations are not random.”
This is based on several mistakes.
– Mutations *are* stochastic (“random”), but they are not necessarily equi-probable (“random”).
Mutations follow stochastic distributions that are chemically constrained. (Different bases have different substitution probabilities; mutation factors are environmentally controlled; et cetera.)
– Selection is indifferent (“orthogonal”) to variation.
Selection results. such as convergence, does not depend on variation as such. Take bioluminiscence as an example, strong purifying selection must conserve luciferin and especially its sequence, despite stochasticity of mutations. On the other hand the luciferases vary, a lot as I understand it, which they can do due to stochasticity of mutations.
“we don’t know how information can acquire causal efficacy in a material substrate”.
Do you refer to emergence, since the process of evolution (rather self fulfilling) predicts the efficiency of having genetics?
Vent theory suggests it came about due to PCR of a molecule (RNA) that happens to be a fair enzyme (among other things) under the same Hadean conditions. If you ask “why” (it is an enzyme et cetera) there is a plethora of possible answers, such as “why not”, but it is a fact rather remote from the process that established efficiency (evolution) as such.
You write “mutations follow stochastic distributions that are chemically constrained” but isn’t reality even stranger than that? As a demonstration, I can’t help thinking of the CAG repeat of HTT gene in Huntington’s disease that, on average, lengthens with each generation. Here we have molecular convergence from many different lines, in the complete absence of any selective advantage.
You mention “overcome notions of the modern synthesis”. This is very scary and almost certainly wrong. Without these notions, the idea started by Darwin is still incomplete and we have nothing that works logically. Sure, you can use non-linearity to try and add higher order concepts on top of it – but not at the expense of destroying the mechanistic basis.
Say you did have such a well formed concept, rather that just nebulous and distracting ideas, well this would give you some idea of what allows evolution to solve some problems. As such you could use the mathematical structure of which your ideas is based to write an algorithm that could do the same on a computer. Given the much faster reproduction time of information in such systems, they could exceed anything biology could produce (such as the human brains ability to play go) in just days. If none of your concepts could ever be used in this fashion, I fear your doing nothing more than invoking vital force.
Note that we can, and have, employed modern synthesis to this task, so far with only limited success. This might indeed suggest another high order concept may help the theory, but, to be of any use, it should be far better formed than the ideas above presented.
Regarding your comment on SETI, you may be very interested in reading this online history of the subject:
http://www.daviddarling.info/encyclopedia/S/SETI_critical_history_cover.html
THIS is why I read and love this blog.
Thankyou.
P
NASA’s idea of a “mission to Europa” isn’t to swim a probe in its ocean.It’s not even to land and test out e.g. thermal boring tech to get down to the ocean. Nope. Their idea is to orbit and pray for a plume. Colour me underwhelmed.
Congress told them to be bolder or else back in May. See my links to these news items in this thread above. Here is one, just in case:
http://www.thespacereview.com/article/3025/1
NASA is very skittish when it comes to finding alien life in the Sol system, even though that is what the public wants them to do more than just about anything else. Such a discovery would no doubt increase their budget considerably. But that is logic, not bureaucracy.
Nasa is probably well aware that the likelihood of finding life in the solar system is remote, and failure to find it after spending a lot might not go down well with funding legislatures, as well as disappointing the public. Their strategy might be considered more of a constant teaser, like “sensational” documentaries on the History Channel that are all about the excitement of the chase, but rarely find anything of consequence.
In the mean time, a lot of good science gets done.
“Nasa is probably well aware that the likelihood of finding life in the solar system is remote, and failure to find it after spending a lot might not go down well with funding legislatures, as well as disappointing the public. ”
Well said and I believe you have well-captured the current thinking at NASA, thinking I agree with.
The posted article is wonderfully written and I have only really skimmed it and will read through again more deeply, as well as some of the truly amazing comments on this thread… but as someone implied in one of the posts in this thread: an ounce of evidence is worth a ton of theory.
We need evidence.
The theories are wonderful but I believe right now, at this juncture, at this point in time in our history as a space-faring race, our best hope for finding evidence of life in our solar system is on Mars. Not that it is likely that there is any life there now, but it is quite conceivable that it was there once.
I vote for my taxes to be spent on Mars. That doesn’t mean saying “no” to other destinations, just that if it comes to a choice between another enhanced Martian Rover and a submarine for Europa… I vote for Mars. (I know, we already have one in the pipeline, Mars 2020. I’m just stating a general preference.)
We have literally only scratched the surface on Mars. And yet we’ve already found solid evidence of ancient water-flows on the red planet.
We need to dig deep, get to the bottom of things on Mars, and I think we’ll uncover solid proof that life evolved there, and that it died out before it had a chance to flourish.
(BTW, viva SJ Gould. He is, for me, the Einstein of Biology.)
Fascinating read.
Regarding:
“First of all, cell types with an increased genomic complexity, eukaryote-like organisms, need to arise. Interestingly, on Earth this transition happened only once (thereby being one of the bona fide evolutionary bottlenecks in the emergence of higher-order biocomplexity; however we cannot exclude that now extinct alternative eukaryote-like cell designs might have existed at some point in Earth’s history).”
Dummy question: should we a-priori give up any hopes of uncovering evidence of this *actually* having occurred in Earth’s history, or could the existence of such alternative eukaryote-like cells have left some sort of signature, detectable by present-day science?
Excellent question, indeed! I think the chances of finding mico-fossils representing such alternate eukaryote-like organizational forms is exceedingly small as our (micro)fossil record is, admittedly, very gappy and incomplete after all.
Luckily, however, there other routes of inferring evolutionary events, i.e. finding “signatures” of possible alternate eukaryotes. Such vestiges would be genomic signatures from extant life forms proving unorthodox evolutionary scenarios in which eukaryotic complexity evolved BEFORE bona fida eubacteria and archaea. I am directing you to an excellent article by EDU colleague Carlos Mariscal detailing these issues. If eukaryote first or a related scenario is actually true, we could infer that early cellular evolution did not proceed according to our common understanding after all, entailing a multitude of eradicated eukaryote-like progenote ancestors.
“In the half century since the formulation of the prokaryote : eukaryote dichotomy, many authors have proposed that the former evolved from something resembling the latter, in defiance of common (and possibly common sense) views. In such ‘eukaryotes first’ (EF) scenarios, the last universal common ancestor is imagined to have possessed significantly many of the complex characteristics of contemporary eukaryotes, as relics of an earlier ‘progenotic’ period or RNA world. Bacteria and Archaea thus must have lost these complex features secondarily, through ‘streamlining’. If the canonical three-domain tree in which Archaea and Eukarya are sisters is accepted, EF entails that Bacteria and Archaea are convergently prokaryotic. We ask what this means and how it might be tested.”
http://carlosmariscal.com/wp-content/uploads/2012/11/20140322.full_.pdf
So, we may eventually find out that even that one seemingly trick step is much less of a bottleneck than our current (2016) understanding would lead us to think. That, in itself, is already so thought-provoking.
Thank you so much, Claudio!
LuigiG
An interesting, but highly speculative article. I’d like to see some proof, rather than philosophy to back up the assertions. I will certainly try to explore some of the referenced papers to understand this better.
Regarding the Enceladus lander experiments. I fail to see how antibody, microarray and Nanopore sequencing can hope to work unless you are assuming that life uses the same chemistry as Earth. If it does, I would suggest this undermines the top down informational argument in favor of a common ancestor. If life is using even slightly different substrates, antibodies might work if you are lucky (but what would a signal mean?), microarray (RNS/DNA) would find nothing, and nanopore technology using pores calibrated for terrestrial DNA will fail to find anything (they will either block or read nothing as different polymers stream through the pore without making electrical contact).
I’m not sure you even need to land, but rather sample a plume instead. All the experiments listed should work on plume material, as long the frozen particles remain large enough to contain cells.
Regarding bioluminescence. The argument rests on the requirements that:
1. bioluminescence evolved to consume oxygen on Earth.
2. that O2 is indeed found in subsurface oceans by peroxide leakage
Both are speculative, but they do fit the Freeman Dyson approach of looking for easy things, so maybe worth a shot.
However, using lights to attract multicellular organisms is a stretch as it assumes they evolved and that they also evolved eyes in a dark world. I would also suggest that horizontal genetic transfer from microbes might be an alternative explanation for the separate evolution of intrinsic bioluminescence in fish. (That should be testable?) If so, that would bolster Lynn Margulis’ hypothesis about some evolved traits.
Hi Alex,
regarding your comments about MCTs and alternative chemistry. I am stating that I am expecting “roughly” the same carbon based chemistry but not necessarily functional biopolymers with molecular identity. Take for example nanopore sequencing and its potential to detect XNAs. Information-storing polymers, according to our knowledge, must be linear and water soluble. Have a look at this paper by my colleague Fabio Rezzonico:
http://www.ncbi.nlm.nih.gov/pubmed/24684166
“Nanopore-based sensing devices allow the analysis of single molecules, including the sequence of informational biopolymers such as DNA or RNA, by measuring current changes across an electrically resistant membrane when the analyte flows through an embedded transmembrane protein or a solid-state nanopore. Under certain basic assumptions about their physical properties, this technology has the potential to discriminate and possibly analyze biopolymers, in particular genetic information carriers, without prior detailed knowledge of their fundamental chemistry and is sufficiently portable to be used for automated analysis in planetary exploration, all of which makes it the ideal candidate for the search for life signatures in remote watery environments such as Mars, Europa, or Enceladus.”
Related to the other instrumentation you mention I am taking a similar broad view as explained for example in:
http://www.hou.usra.edu/meetings/lpsc2014/pdf/2653.pdf
and
http://www.ncbi.nlm.nih.gov/pubmed/24735484
“We make a specific recommendation that certain characteristic patterns in the composition of lipid hydrocarbons can only result from a biological process, because the signal arises from a universal requirement related to lipid bilayer fluidity and membrane stability. Furthermore, the pattern can be preserved over millions of years, and instrumentation is already available to be incorporated into flight missions.”
In response to your doubts about the emergence of bioluminescence in multicellular organisms, have look at these articles that I also linked in the essay:
https://www.genome.gov/27551984/2012-news-feature-bioluminescent-comb-jellies-begin-to-shed-light-on-the-evolution-of-vision/
http://phenomena.nationalgeographic.com/2016/06/08/the-many-origins-of-glowing-fish/
Best regards,
Claudio
The first thing which came to my mind was “Evolution: A View from the 21st Century”, 2011, James A. Shapiro, and his theorem of natural genetic engineering. On a second thought i feel reminded about Lovelock’s Gaia hypothesis. A paradigm change of some sort is in order, i guess. Both were framed for creationism, tho, so be careful.
I have to admit i always had trouble with convergent evolution as a concept, to be honest. The mathematical improbabilities just seemed too abysmal to me and, as i realized whilst looking at horizontal gene transfer, for good reasons. I am wondering if such evolutionary achivements were really achived as independantly as it is currently concieved.
Biomolecular information proccessing is reality. There is guiding logic behind this even if it hardly equals to what is currently understood as sentience. Its as different a concept as it is imaginable, hence its logic is guided by different principles. Yet, obviously its in comparision far more advanced with respect to promblem solving than we ourselves are.
It makes one wonder with respect to such events as the (possibly so called) oxygen catastrophe some some 2,3 billion years ago, paving the way for animal life on the planet as a “solution” are really that coincidental or actually a case of environmental engineering. Its creepy to think about it. I am generally suspicious when it comes to coincidences, especially of the planet altering type.
This line of thinking opens up a totally different can of worms with respect to abiogenesis on this planet alltogether. Hawking put it this way: it is surprising that such a complex coincidence like the emergence of life on Earth happened so early during the planetary habilitable period.
Indeed. Another planet altering coincidence. And even more abysmal improbabilities to overcome with respect to translation and transcription machinery.
Taking into account our chronological record of life’s history in our sediments as well what we can deduce from genetics (LUCA) some episodes in evolution seem to happen unusually quickly and those are especially the most complex ones (protocellular to cellular, cambrian explosion, etc).
There is no question about the evolution of life on this planet, however it makes one wonder, especially with respect to environmental conditions during the hadean eon and the general delicacy of an emerging replicating process with no initial evolutionary memory to fall back onto for adaptation to these circumstances if life really emerged on this planet.
Its also certainly one of the pitfalls we will encounter, should we succeed in finding extraterrestrial life in the solar system and it is similar to our own. The question, considering overall microbial spaceworthiness, will be asked if it indeed was a case of convergent evolution or maybe a sort of seeded contamination.
Probability wise i had to settle on the latter for that respect, i am afraid. I also get the impression this is only the tip of the iceberg. We’ll see.
“The emergence of intelligence is also obviously not “contingent” as neurons have evolved multiple times in animals.”
1) There’s the 2014 Moroz paper, which is certainly very interesting and provocative, suggesting an independent evolution for neurons in Ctenophora. However, my impression is that there is not yet a clear consensus on this point. Moroz has made a plausible prima facie case, but much more research needs to be done. The Moroz paper itself hedges its bets, and points to several areas that need closer examination — “weak support is probably due to under-representation of comparable transcriptomes from sponges and large protein divergence,” usw. He also notes that it’s still conceivable that Ctenophora could be a clade within Eumatazoa — just that this is “less parsimonious”, since it would require the ctenophores to lose a lot of their molecular machinery at some point, and then evolve it all over again it in a different way. That’s a fair point, but on the other hand sometimes evolution works that way — cetaceans have horizontal tail-flukes instead of vertical fins, and so forth.
Anyway: if there’s now a general put-it-in-the-textbooks consensus that Moroz et al. are correct, I’d be interested to hear.
2) Twice is not “multiple times”.
3) Is it really all that surprising that neurons (and possibly muscles) might have evolved twice? Especially given that the innovation would have occurred in both groups right around the same time, in the late Neoproterozoic.? Available evidence suggests that this was an extremely exciting time in animal evolutionary history — in some ways, more interesting than the famous Cambrian period that followed immediately after. Even if neurons did evolve twice, it happened only during that particular odd period. They never evolved before, or since. Perhaps there was simply a one-time paleoenvironmental “window” that encouraged this particular adaptation in two separate metazoan groups.
4) Neither neurons nor close analogs of neurons have evolved in any other multicellular group outside the Animalia. An oak tree is a massive, morphologically complex eukaryote with dozens of different specialized tissue types, but it does just fine without neurons. “Evolved twice within the Animalia” does not seem terribly compelling as a universal principle unless you can demonstrate that the Animalia — or something like them — are a universally probable outcome.
As far as we can tell, given limited current knowledge, the Animalia don’t seem to have started diverging from other opisthokonts until quite late in the Neoproterozoic — over a billion years after the GOE, and probably several hundred million years after the emergence of the first eukaryote. Perhaps their eventual emergence was “not contingent”, but it doesn’t look like evolution was in any hurry here.
Doug M.
Hi Doug,
thanks for your feedback.
1) I agree, these issues are not conclusively proven yet. More comprehensive phylogenomic taxon sampling is needed to settle these issues. Then again, the phylogenetic after major transitions, i.e. in this case the emergence of animal complexity, always tends to be noisy, ambiguous and even paradoxical.
2) Neurons probably evolved more than twice in MULTIPLE cell lineages even within individual phyla. According to my working hypothesis at least three times in Ctenophora, Cnidaria and Bilateria. Possibly many more times within Bilateria. You can wait for my upcoming paper on this interesting problem, I think it will add some novel arguments to the debate.
3) According to neo-Darwinism, yes, it is a really extraordinary event. According to CCE, it’s not extraordinary but expected. I will refer you to this excellent, recent paper by Liebeskind summarizing the convergence perspective:
http://www.cell.com/trends/ecology-evolution/abstract/S0169-5347(15)00302-X
4) This is not correct, there is increasing evidence for cognition in aneural contexts, for example in plants and unicellular eukaryotes, even bacteria. Some instantiations of the underlying functional systems are almost synaptic in their organization. Here I am pointing you to the following trailblazing papers:
http://www.tandfonline.com/doi/abs/10.1080/03081079.2014.920999
http://journal.frontiersin.org/article/10.3389/fpsyg.2015.01329/full
Thanks again for your insightful remarks.
Best,
Claudio
It is always interesting to see someone work into a science career. Having hands on experience in some form (Enceladus mission proposals) should help!
Being interested in astrobiology, currently studying Bioinformatics at a prestigious university, and in fact having a (non-biology) PhD, could put me in a position to rudimentary criticize the article. Since it is attacking the consensus theory of the field, the author is surely prepared on harsh such. The article is covering a lot, so for brevity I will omit most references this time around.
A minor criticism is the inflated format and use of philosophy, which I gather has historical roots in biology. [Yeah, not a biologist!] YMMV; personally, with roots in physics, I find it detracts from communication and understanding. Astrobiology is greater than biology.
Also, the ideas of evolutionary convergence and emergence as contributing putative new biological laws have references in Conway Morris, who are a theist, and Davies, who are a deist. That raises flags.
Now to the meat:
1. The modern synthesis [MS], aka neo-darwinism in philosophical terms. is alive and well. There isn’t any viable alternative.
2. The article explores a lot of possible MS alternatives, which is popular but doesn’t test the alternatives as such.
3. A universal biology wouldn’t be expected to have universal laws except the existence of the biological evolutionary process itself. Conversely, the conjured up “replication cycles” of cosmological biological components seems both lacking in examples and other mechanisms, since evolution is a process within populations of a biosphere.
4. Evolutionary convergence is not a generic outcome. It may happen from divergent genotype under strong constraints (say, the fusiform body shape of fast swimmers in dense media) or from same genotype under loose constraint (say, the bioluminiscence examples given that are all based on extremely preserved luciferin).
Re contingency as an outcome, the challenge to necessarily incorporate life emergence under evolution seems a mistake. Those are separate processes (geology respectively biology) even though they may share mechanisms. And while contingencies of traits like language capable intelligence seem rare, the short time to emergence of life implies it isn’t a contingent outcome of geology.
5. MCT incorporates phenotype that is generally given. We wouldn’t call the spherical form of water droplets or sufficiently large planets a result of biological convergence. Conversely, cell forms, membranes and metabolic like pathways of life seems to have been present where life emerged. (Weiss et al recent result on the LUCA testing the vent theory.) PCR of replicator molecules have also been hypothesized and lab tested under the same theory.
This may be a test of MCT; I certainly think it has failed and need convincing otherwise.
6. Emergency of physics which is in principle impossible to derive from the underlying template is consistent with physics (say, electron shell structure of molecules), emergence of life (say, plate tectonics) and biology (say, quantum mechanisms of antenna structures in photosynthesis).
7. Information or architectural transitions have never played an important role in evolution theory as such.
As notes here eukaryotes split off within Archaea. [By a result of Spang from Uppsala University, which is both my alma mater and where I study Bioinformatics, by the way.] They have shown to evolve multicellularity on time scales of a year in the lab. But prokaryotes thrive in evolving it too, some 20 times. Mostly as fruiting bodies, but cyanobacteria arguably evolved complex multicellularity once. (Differentiated heterocysts would constitute soma, since they don’t participölate in the germ line.)
8. Epigenetics is not based on non-DNA information but encoded for transcription factors.
Henikoff, epigenetic expert:
“… transcription factors occupy the top of the hierarchy of epigenetic information, [that] this has been widely accepted in the broader chromatin field, and [that] histone modifications at most act as cogs in the machinery that enforces the often complex programs specified by the binding of transcription factors. In no case that I recall is there an example of a change in gene expression that can be attributed to histone hyperacetylation to the exclusion of non-histone substrates, of which many have been identified.”
[ https://whyevolutionistrue.wordpress.com/2016/05/05/the-new-yorker-screws-up-big-time-with-science-researchers-criticize-the-mukherjee-piece-on-epigenetics/ ]
Minor problems:
1. Confusing the sequence hypothesis of Watson (protein-RNA-DNA) with the central ‘dogma’ of Crick (once sequence information gets into protein it doesn’t get out). [A good review, I remember the textbook in a molecular biology course I took decades ago cemented the mistake: http://sandwalk.blogspot.se/2007/01/central-dogma-of-molecular-biology.html ]
2. Bacteria and Archaea evolved from a lipid membrane LUCA. [See the Weiss et al result.] The modern membranes are divergent, not convergent.
3. Consensus on ctenophores are that they split after sponges did. The results that implied otherwise went away with more sequencing. But even if not, evolution of neurons twice would not tear down MS.
I am sure i missed a lot in such a long article, but hopefully not the major points of it. And that I made a lot of mistakes since my comment is large and somewhat detailed (and I confess I didn’t check all the available literature). But criticism of criticism is both welcome and fair dinkum science.
So I noted a mistake of mine myself. I inverted the sequence hypothesis but didn’t notice: DNA-RNA-protein.
Hi Torbjörn,
thanks for your comprehensive analysis of my essay, it took a while to answer. Sorry for that!
“A minor criticism is the inflated format and use of philosophy, which I gather has historical roots in biology. [Yeah, not a biologist!] YMMV; personally, with roots in physics, I find it detracts from communication and understanding. Astrobiology is greater than biology.”
I didn’t know that physics is an ahistorical universal system of knowledge that exists independently of philosophical enquiry. But wait…doesn’t modern physics have its roots in META-physics?
“Also, the ideas of evolutionary convergence and emergence as contributing putative new biological laws have references in Conway Morris, who are a theist, and Davies, who are a deist. That raises flags.”
This doesn’t add anything to the issues debated. Darwin, for exampe, was greatly inspired by the natural mysticism that his greatfather Erasmus preached. I could spin this around by saying “Oh Darwinism is a big conspiracy, because Darwin was a freemason”. No scientific argument here.
“1. The modern synthesis [MS], aka neo-darwinism in philosophical terms. is alive and well. There isn’t any viable alternative.
2. The article explores a lot of possible MS alternatives, which is popular but doesn’t test the alternatives as such.”
Clearly, this is a circular argument. There are MANY alternatives, some more radical than others, I prefer the more radical ones (following proposals figure as the “extended” synthesis, I am calling for a novel even more encompassing “integrated” synthesis):
https://en.wikipedia.org/wiki/Extended_Evolutionary_Synthesis
http://www.thethirdwayofevolution.com/
http://www.nature.com/news/does-evolutionary-theory-need-a-rethink-1.16080
“3. A universal biology wouldn’t be expected to have universal laws except the existence of the biological evolutionary process itself. Conversely, the conjured up “replication cycles” of cosmological biological components seems both lacking in examples and other mechanisms, since evolution is a process within populations of a biosphere.”
Your unprovable metaphysical assertion is that the Universe is an expanding dead block of matter including biological molecular structures that magically act in complex ways but are nothing more than the dead matter they are composed of. I beg to differ, in that I am proposing that just as we are alive, the Universe is alive.
“4. Evolutionary convergence is not a generic outcome. It may happen from divergent genotype under strong constraints (say, the fusiform body shape of fast swimmers in dense media) or from same genotype under loose constraint (say, the bioluminiscence examples given that are all based on extremely preserved luciferin).”
I agree, although I don’t know what exactly is being critized here. CCE is encompassing the origin of biophilic universes that render the emergence of life very likely or – almost – necessary. From galactic, solar, planetary, geological and biological evolution. It’s a continuum. The super-entity itself is organic and alive, although some sub-structures contained therein are certainly not “alive” (but are part of the organic whole). I am not arguing for somekind of animism.
“5. MCT incorporates phenotype that is generally given. We wouldn’t call the spherical form of water droplets or sufficiently large planets a result of biological convergence. Conversely, cell forms, membranes and metabolic like pathways of life seems to have been present where life emerged. (Weiss et al recent result on the LUCA testing the vent theory.) PCR of replicator molecules have also been hypothesized and lab tested under the same theory.
This may be a test of MCT; I certainly think it has failed and need convincing otherwise.”
Unfortunately, I don’t really get the point here, maybe you misunderstood the notion of MCTs. MCTs are also not a result of (pure) biological convergence but rather cosmic convergent evolutionary processes.
“6. Emergency of physics which is in principle impossible to derive from the underlying template is consistent with physics (say, electron shell structure of molecules), emergence of life (say, plate tectonics) and biology (say, quantum mechanisms of antenna structures in photosynthesis).”
Since philsophy is anathema in science, I am going to obey.
“7. Information or architectural transitions have never played an important role in evolution theory as such.”
You should have a look at the papers I cited before negating my arguments without even adressing the issues at hand. Have a look at the the classic “The Major Transitions in Evolution”, it’s all about informational architecture.
“8. Epigenetics is not based on non-DNA information but encoded for transcription factors. ”
This is a very limited if not plain out wrong view on epigenetics. I worked in two different epigenetics labs and the consensus view on epigenetics was certainly much broader than yours, more according to:
http://www.ncbi.nlm.nih.gov/pubmed/23691975
In summary, I think you are fully entitled to your view on evolution and my essay. On other hand, it won’t take much more time, maybe 10 years, and the limits of the MS will be mainstream knowledge, thereby leading to the more comprehensive paradigm that Woese demands. By definition, these theories are only models and do not have absolute reality. Reductive materialists just tend to feel insulted if someone opines that there might be more to biology and human existence than pure chance and a cosmic abyss of meaninglessness. Proposing that there is telos in nature doesn’t mean that there is a supernatural force responsible for the emergence of life. It just means life generates life.
Best,
Claudio
Using “long range ultrasonic sounds” to detect living/non-living objects might be a good idea, if the target moved and behaved like a fish then it could be a fish (it’s not like I believe there are fishes living in the ocean of Europa anyway, this might be a dumb example from an ignorance).
Second, technological convergence might or might not be able to happen. It’s well known there are many people who like to build Dyson Sphere or Von Neumann Machines for colonizing the whole galaxy etc… However, these are not the optimal ways to move up + forward to become an elder civilization in far future, there are some concepts which are much much more interesting to attain than those mentioned above.
As a sidenote, i really like to see these subs exploring our interplanetary oceans. However, sampling material, maybe even a sample return mission by flying through ejecta, as the Cassini mission did at Enceladus is so much simple. I guess such missions would be a good idea to select targets before sending the subs. Bioluminescence as a biomarker is a nice idea, however there can also be geologic sources of light, especially with high tectonic activit as one would expect from one of Jupiter’s moons. That being said, its not to difficult to tell one from the other with a bit of thermal sensory equipment.
Flying through a plume at Enceladus and using aerogel to collect the materials makes a lot of sense, but it may not work for Europa since the initial reports of geysers have not produced further detections since:
https://centauri-dreams.org/?p=32148
And are there volcanoes on Titan? I know there are active volcanoes all over Venus: Perhaps an aerostat mission to sample the atmosphere then send back samples via rocket.
I second Phil’s comment! An amazing, sprawling, thought provoking piece that connects Chalmer’s thinking about consciousness to the GHZ.
Paul, thanks so much for your wide ranging net.
Well this news is ironic and not in a good way, in that Congress wants a full-on mission to Europa and NASA is the one that seems to be hedging. See here:
Europa Clipper Team Braces For Bad News
Published: 19 August 2016
by Evan Gough
Jupiter’s moon Europa is a juicy target for exploration. Beneath its surface of ice there’s a warm salty, ocean. Or potentially, at least. And if Earth is our guide, wherever you find a warm, salty, ocean, you find life. But finding it requires a dedicated, and unique, mission.
If each of the bodies in our Solar System weren’t so different from each other, we could just have one or two types of missions. Things would be much easier, but also much more boring. But Europa isn’t boring, and it won’t be easy to explore. Exploring it will require a complex, custom mission. That means expensive.
NASA’s proposed mission to Europa is called the Europa Clipper. It’s been in the works for a few years now. But as the mission takes shape, and as the science gets worked out, a parallel process of budget wrangling is also ongoing. And as reported by SpaceNews.com there could be bad news incoming for the first-ever mission to Europa.
Full article here:
http://www.universetoday.com/130357/europa-clipper-team-braces-for-bad-news/
To quote:
What all this seems to mean is that the initial science and instrumentation for the mission will be maintained, but no additional capacity will be added. NASA is no longer considering things like free-flying probes to measure the plumes of water ice coming off the moon. According to Niebur, “The additional science value provided by these additions was not commensurate with the associated impact to resources, to accommodation, to cost. There just wasn’t enough science there to balance that out.”
If you are an American citizen reading this and I presume care about exploring space, let NASA know that not exploring Europa is more than just short-sighted, it is an insult to scientific progress, especially exobiology. If you are not an American citizen but want to see a full-fledged Europa mission happen, let NASA know just the same.
Another quote:
“It’s unfortunate that a few tens of million dollars here or there can hamper our search for life beyond Earth. But the USA is a democracy, so that’s the way it is. These discrepancies and possible disputes between NASA and the different levels of government may seem disconcerting, but that’s the way these things get done.”
That’s just the way it is? Really? Science by popular vote? I don’t think so.
Contact NASA. Contact your political representative. Contact some scientists and tell them to use their clout to make this happen. Maybe Neil deGrasse Tyson can put his fame to good use.
Well… budget is not infinite. And they have other high priority mission goals at this time. Orion. Manned flight to Mars. Its the old manned mission vs getting the most bang for science, which is robotic exploration currently. National pride plays a huge part, especially with having to rely on Roscosmos for astronaut transport and of course the simple truth that we WANT to get humans into space. Then there is the Webb telescope, wich i personally consider an excellet choice. The question is what and how fast. Personally i’d take a huge chunk out of defence budget and put it into NASA budget, which is a relatively tiny, tiny fraction of the overall expensees. In short: science is, as usual undervalued and military, as usual overvalued. The kinds of fraud affairs we know from military spending are unheard of from NASA projects, tho accidents do happen of course, as well. There are of course JAXXA, ESA, Roscosmos, CSNA and even ISRO. All in all progress is a bit slow currently but i’ll even out eventually. It would be nice if there would be a global agency with funding of serveral leading nations and generally more focus on space exploration globally. As much as you can complain about the NASA budget, its NASA which has accomplished the bulk of exploration work internationally. So, what about other countries? You can’t always put this squarely on American shoulders. Why is there so little spending for exploration and so much for warfare? This is totally skewed twoards a detrimental outcome. Hasn’t to be like this.
Is anyone thought of any contingency of whether or not what might happen to the probe, if the creature encounters decides to take a bite out of it or even eat the thing? Not trying to be funny here. What makes you think that a large creature couldn’t possibly think this thing was a snack?
It would mean we have discovered life on another world. I was at a lecture on the 2004 Mars Rovers when a child asked the speaker what would happen if a Martian ate a rover. He said it would be a great discovery. :^)
NASA’s Europa Mission Facing Possible Budget Cuts in 2017
By Paul Scott Anderson
For a long time now, there has been growing interest in sending a mission back to Jupiter to better study one moon in particular: Europa. Previous missions such as Voyager and Galileo showed us this world up close for the first time, revealing a place that maybe, just maybe, is home to some kind of life.
On the outside, Europa is cold and frozen, like an airless version of Antarctica, with its surface completely composed of ice [and lots of surrounding nasty radiation from Jupiter.]. But deeper down, as those probes found, there is a global ocean of water deeper than any oceans on Earth. In more recent years and months, a new NASA mission to Europa has finally started to take shape, with a launch tentatively scheduled for 2022. As often happens, however, the mission is facing possible budget cuts in 2017.
As reported by SpaceNews, the mission planning and development is still continuing, but there may be a squeeze in funding for the next fiscal year.
“There is this squeeze in FY17 that we have,” said Bob Pappalardo, the project scientist at the Jet Propulsion Laboratory. “We’re asking the instrument teams and various other aspects of the project, given that squeeze, what will it take in the out years to maintain that ’22 launch. We’re actively pursuing that question as we speak.”
During presentations at NASA’s Outer Planets Assessment Group (OPAG) in Flagstaff, Ariz., on Aug. 11, it was announced that NASA is looking for ways to cut costs in 2017, but also keep the mission on track for a launch in 2022.
Full article here:
http://www.americaspace.com/?p=94964
To quote:
The overall goal of a Europa mission is, of course, to look for possible evidence of life. The Europan ocean would be dark and cold, but even in similar environments on Earth, life flourishes. As Niebur noted: “Mission success, in NASA’s mind for this mission, which is 23 or so days long, is that on day 25 we have a press conference at NASA Headquarters and announce that we’ve found life. If that is an unrealistic expectation, I need to know it now and not on day 22 of the mission.”
No reason cited why Nasa requested such a small sum, nor why the much larger sum granted by Congress for FY2017 cannot be assumed to be operative. Is this Nasa playing internal politics or trying to shift funding around to support other projects?
Probably both. My concern is that they have too many fingers in too many pies and not enough money to go around. $18 billion annual budget is chicken feed compared to most other government agencies.
One year of funding for the Department of Defense (DoD) is currently equivalent to three decades of annual NASA budgets. Their spending waste alone costs more than the entire space agency several times over. We could launch a dozen probes to Europa on that alone.
Imagine what NASA could accomplish – and succeed with – if they had $400 billion to work with:
http://www.cnn.com/2016/08/26/politics/f-35-fighter-jet-problems-gilmore-memo/index.html
The Future Planetary Exploration blog has a new detailed post about the finalists in NASA’s newest New Frontiers program:
http://futureplanets.blogspot.com/2016/08/selecting-next-new-frontiers-mission.html
With an allocated budget of about one billion dollars, there aren’t any NF Europa missions, but check out these two candidates:
* Saturn Atmospheric Probe – Determine the composition of Saturn’s atmosphere to help us better understand the formation of the solar system
* Ocean Worlds (Titan and/or Enceladus) – Do these two moons of Saturn have the conditions to support life and is life present?
To quote:
The first five of these themes were selected through the Decadal Survey. NASA’s managers added the Ocean Worlds theme in response to a Congressional directive and further discoveries by the Cassini mission. For the next, fifth New Frontiers competition, Jupiter’s moon Io and a lunar geophysical network theme will be added.
and…
I suspect that for many readers of this blog, a mission to return to Enceladus or Titan to continue their exploration with a new generation of spacecraft and instruments would be a personal favorite. I share that desire, but also recognize the challenges any proposal to these worlds would face. First, these worlds were just added to the list of candidate themes in the past few months. The in-depth analysis of objectives for these missions is just getting underway by the scientific community. Second, the technical maturity of instruments to explore their oceans, determine their habitability, and search for life may be low – NASA has not made major investments in these technologies for these worlds (but plans to begin to do so). And third, these missions are likely to need radioisotope power generators and their cost would eat significantly into the mission budget, potentially making it less competitive. (Solar powered missions are possible at Saturn, but appear to be on the edge technically. This could make a proposal that depends on solar power appear technically risky.) Balancing these negatives is a heritage of three Discovery-class proposals to these worlds that were not selected but which could form the basis of a New Frontiers-class mission. Still, I personally doubt that a mission to these moons will be selected this time. (If I am wrong, given a mid-2020’s launch and a flight that could last 10 years, it could be the mid-2030s before the spacecraft arrives at its target.)
and…
The Ocean Worlds mission theme is focused on the search for signs of extant life and/or characterizing the potential habitability of Titan and/or Enceladus. For Enceladus, the science objectives (listed without priority) of this mission theme are:
• Assess the habitability of Enceladus’ ocean; and
• Search for signs of biosignatures and/or evidence of extant life.
For Titan, the science objectives (listed without priority) of the Ocean Worlds mission theme are:
• Understand the organic and methanogenic cycle on Titan, especially as it relates to prebiotic chemistry; and
• Investigate the subsurface ocean and/or liquid reservoirs, particularly their evolution and possible interaction with the surface.
The Saturn Probe mission theme is intended to deploy one or more probes into Saturn’s atmosphere to directly determine the structure of the atmosphere as well as noble gas abundances and isotopic ratios of hydrogen, carbon, nitrogen, and oxygen. The science objectives (listed without priority) of this mission theme are:
• Determine noble gas abundances and isotopic ratios of hydrogen, carbon, nitrogen, and oxygen in Saturn’s atmosphere; and
• Determine the atmospheric structure at the probe descent location.
https://arxiv.org/abs/1608.06930
Biofluorescent Worlds: Biological fluorescence as a temporal biosignature for flare star worlds
Jack T. O’Malley-James, Lisa Kaltenegger
(Submitted on 24 Aug 2016)
Habitability for planets orbiting active stars has been questioned. Especially, planets in the Habitable Zone (HZ) of M-stars, like our closest star Proxima Centauri, experience temporal high-ultraviolet (UV) radiation. The high fraction of M-stars (75%) within the solar neighborhood, the high occurrence rate of rocky planets around M-stars, and the favorable contrast ratio between the star and a potentially habitable rocky planet, makes such planets interesting targets for upcoming observations.
During M-star flares, the UV flux on a HZ planet can increase by up to two orders of magnitude. High UV radiation is harmful to life and can cause cell and DNA damage. Common UV protection methods (e.g. living underground, or underwater) would make a biosphere harder to detect. However, photoprotective biofluorescence, “up-shifting” UV to longer, safer wavelengths (a proposed UV protection mechanism for some corals), would increase the detectability of biota and even uncover normally hidden biospheres during a flare. Such biofluorescence could be observable as a “temporal biosignature” for planets around UV-active stars.
We model temporal biofluorescence as a biosignature for an exoplanet biosphere exposed to such conditions, based on planets in M-star HZs. We use fluorescing coral proteins to model biofluorescence, comparing observable spectra, and colors, to vegetation and fluorescent minerals. Our planetary models assume a present-day Earth atmosphere and explore the effect of varying cloud coverage and land:ocean fractions.
UV flare-induced biofluorescence could be remotely detectable, comparable in strength to vegetation on Earth. On planets in the HZ of M-stars, biofluorescence could be a temporary biosignature, distinguishable from fluorescing minerals and vegetation.
Comments: Submitted to Ap.J., 16 pages, 10 figures
Subjects: Earth and Planetary Astrophysics (astro-ph.EP)
Cite as: arXiv:1608.06930 [astro-ph.EP]
(or arXiv:1608.06930v1 [astro-ph.EP] for this version)
Submission history
From: Jack O’Malley-James [view email]
[v1] Wed, 24 Aug 2016 19:53:19 GMT (5895kb)
https://arxiv.org/ftp/arxiv/papers/1608/1608.06930.pdf
Thanks ljk,
very interesting article. I presented my hypothesis on bioluminescence and convergent biosignatures during a lecture of Prof. Kaltenegger when she was still working in Heidelberg. This seems to be inspired by this:
“Corals were one of the the earliest forms of animal life to evolve on Earth, with an evolutionary history spanning 500 Myr. Initially existing as simple, solitary organisms, they later evolved into collective reefs. Reef-building is an ancient trait for life on Earth. Many marine species have independently evolved reefbuilding abilities, but the trait can be traced back to colonial groups of bacteria building stromatolites 3.5 Gya (Kiessling 2009). The fluorescent proteins in corals are descended from green fluorescent proteins (Field et al. 2006). Green fluorescent proteins are present in a variety of phyla, suggesting an origin within an ancient common ancestor in the very early metazoan (animal) life, over 500 Myr ago (Chudakov et al. 2010). Biofluorescence is widespread in life on Earth and is thought to have evolved independently, multiple times (Sparks et al., 2014; Gruber et al. 2015; Gruber & Sparks, 2015), which strengthens a case for the evolution of biofluorescence on other inhabited worlds, following convergent evolution arguments.”
Bioluminescence and biofluorescence are closely related physical processes. The whole idea idea fits the CCE paradigm.
Best,
Claudio
Very glad that this article was of use to you, Claudio. Now let us hope the first submersible Europa explorer is not only equipped to search for bioluminescence but also has a series of external lights it can control in order to attract any aquatic Europan natives – and perhaps communicate with as well, since I am already in deep waters.
https://arxiv.org/abs/1608.07372
The surface temperature of Europa
Yosef Ashkenazy
(Submitted on 26 Aug 2016)
Previous estimates of the surface temperature of Jupiter’s moon, Europa, neglected the effect of the eccentricity of Jupiter’s orbit around the Sun, the effect of the eclipse of Europa (i.e., the relative time that Europa is within the shadow of Jupiter), and the effect of Europa’s internal heating.
Here we estimate the surface temperature of Europa, when Europa’s obliquity, eclipse and internal heating, as well as the eccentricity of Jupiter, are all taken into account. For a typical internal heating rate of 0.05 W/m2 (corresponding to an ice thickness of about 10 kms), the equator, pole, and global mean surface temperatures are 101.7 K, 45.26 K, and 94.75 K, respectively.
We found that the temperature at the high latitudes is significantly affected by the internal heating. We also studied the effect of the internal heating on the mean thickness of Europa’s icy shell and conclude that the polar region temperature can be used to constrain the internal heating and the depth of the ice. Our approach and formalism can be implemented to estimate the temperature of other moons in the solar system.
Subjects: Earth and Planetary Astrophysics (astro-ph.EP)
Cite as: arXiv:1608.07372 [astro-ph.EP]
(or arXiv:1608.07372v1 [astro-ph.EP] for this version)
Submission history
From: Yosef Ashkenazy [view email]
[v1] Fri, 26 Aug 2016 07:05:05 GMT (84kb,D)
https://arxiv.org/pdf/1608.07372v1.pdf
Russia wants to land a probe on nearby Ganymede:
http://www.dailygalaxy.com/my_weblog/2016/09/russia-announces-space-mission-to-land-a-rocket-on-jupiters-ganymede-largest-moon-in-our-solar-syste.html
I might feel a bit more confident with their claim if they could a probe working past Mars, which they have yet to do. And as for Mars, they haven’t had a successful mission to the Red Planet since Mars 5 in 1974 and the partially successful Phobos 2 in 1988, both during the Soviet era.
See this superb monograph about Soviet Mars exploration:
http://history.nasa.gov/monograph15.pdf
Review: All These Worlds Are Yours
The search for life beyond Earth has attracted a lot of public interest, but where is the best place to look for such life? Jeff Foust reviews a book that offers an astrobiological survey of the solar system and beyond.
http://www.thespacereview.com/article/3059/1
To quote:
Throughout the discussion in the book, Willis pits the different destinations and approaches against one another, in a sense: he asks the reader how they would spend a hypothetical $4 billion astrobiology budget (an amount, he said, he chose because it’s the amount spent globally each day on defense) on ways that might best answer the question of if and where life exists beyond Earth. Do you go all-in on Mars or Europa, for example, or make more investments on exoplanet studies? Willis discloses at the end of the book he would prefer spending much of that $4 billion on a mission to Enceladus, collecting samples of its subsurface ocean expelled into space by plumes for return to Earth. But the message is clear: the search for life requires tough choices given the likely resources available for the effort.
There’s another message in All These Worlds Are Yours as well: finding life beyond Earth won’t be easy. “Basic life will not wave to our passing spacecraft and peering telescopes,” he writes. “Instead, we will be offered mysteries, anomalies, things that don’t add up.” It’s worth adding that some of those mysteries and anomalies may be evidence of life, others simply curious phenomena that have nothing to do with life, and yet others instrument error or interference. That’s a lesson worth remembering the next time you read about the discovery of an interesting alien signal or an Earth-like exoplanet.
http://www.nasa.gov/press-release/nasa-to-hold-media-call-on-evidence-of-surprising-activity-on-europa
Sept. 20, 2016
MEDIA ADVISORY M16-111
NASA to Hold Media Call on Evidence of Surprising Activity on Europa
NASA will host a teleconference at 2 p.m. EDT Monday, Sept. 26, to present new findings from images captured by the agency’s Hubble Space Telescope of Jupiter’s icy moon, Europa.
Astronomers will present results from a unique Europa observing campaign that resulted in surprising evidence of activity that may be related to the presence of a subsurface ocean on Europa. Participants in the teleconference will be:
Paul Hertz, director of the Astrophysics Division at NASA Headquarters in Washington
William Sparks, astronomer with the Space Telescope Science Institute in Baltimore
Britney Schmidt, assistant professor at the School of Earth and Atmospheric Sciences at Georgia Institute of Technology in Atlanta
Jennifer Wiseman, senior Hubble project scientist at NASA’s Goddard Space Flight Center in Greenbelt, Maryland
To participate by phone, media must contact Dwayne Brown at 202-358-1726 or dwayne.c.brown@nasa.gov and provide their media affiliation no later than noon Monday.
Audio of the teleconference will stream live on NASA’s website at:
http://www.nasa.gov/live
For information about NASA’s Hubble Space Telescope, visit:
http://www.nasa.gov/hubble
The Green Universe: A Vision
Freeman Dyson
October 13, 2016 Issue
An illustration of Freeman Dyson’s vision of ‘Noah’s Ark culture’—a space operation in which, ‘sometime in the next few hundred years, biotechnology will have advanced to the point where we can design and breed entire ecologies of living creatures adapted to survive in remote places away from Earth.’ Spacecraft resembling ostrich eggs will bring ‘living seeds with genetic instructions’ to planets, moons, and other ‘suitable places where life could take root.’ A new species of warm-blooded plants, ‘kept warm by sunlight or starlight concentrated onto it by mirrors outside,’ will enable the Noah’s Ark communities to survive.
Full article here:
http://www.nybooks.com/articles/2016/10/13/green-universe-a-vision/
To quote:
Such a proposal would make sense as a first step in a continuing sustained program of exploration of Enceladus. It makes no sense as an isolated one-shot venture. It unfortunately belongs to the NASA Big Space culture, the same culture that gave us the Viking mission to Mars in 1975. Viking was also a one-shot venture, announced with great fanfare as giving a decisive answer to the question whether there is life on Mars. When Viking found no evidence of life, the further exploration of Mars was abandoned for twenty years.
The effect of the Enceladus sample return mission, if it were a one-shot venture like Viking, would probably be the same. Even if kelp is sprouting and sharks are swimming in the Enceladus ocean, the spattered droplets collected from its geysers would probably show no conclusive evidence of life, and the essential question would remain unanswered. The most likely result of a sample return mission would be to raise new questions for following missions to answer. To discover life on an unexplored world will never be a job for a single mission.
and …
All three books look at the future of space as a problem of engineering. That is why their vision of the future is unexciting. They see the future as a continuation of the present-day space cultures. In their view, unmanned missions will continue to explore the universe with orbiters and landers, and manned missions will continue to be sporting events with transient public support. Neither the unmanned nor the manned missions are seen as changing the course of history in any fundamental way.
The authors are blind to the vision of Konstantin Tsiolkovsky, the prophet who started thinking seriously about space 150 years ago. Tsiolkovsky saw the future of space as a problem of biology rather than as a problem of engineering. He worked out the theory of rockets and saw that rockets would solve the problem of space travel, to get from here to there. Getting from here to there is the problem of engineering that Tsiolkovsky knew how to solve. That is the easy part. The hard part is knowing what to do when you have got there. That is the problem of biology, to find ways to survive and build communities in space, to adapt the structures of living creatures, human and nonhuman, so they can take root in strange environments wherever they happen to be. Tsiolkovsky knew nothing of biotechnology, but he understood the problems that biotechnology would enable us to solve.
Yup, it’s water plumes:
http://www.nasa.gov/press-release/nasa-s-hubble-spots-possible-water-plumes-erupting-on-jupiters-moon-europa
Another discovery of water in space – how boring. We must search for alien life
The latest find on Jupiter’s moon Europa offers the potential to really look for life. It’s not time to shy away – and Nasa needs to rise to the challenge
Stuart Clark
Wednesday 28 September 2016 07.20 EDT
For a decade now, the Nasa mantra for looking for life in outer space is to “follow the water”. In the beginning, this was a valid way to identify the places where we should then start searching. Each new discovery of “water” is automatically paired with a statement that this increases the chance of finding alien life. But in recent years – the latest example being the discovery of water this week on Jupiter’s moon Europa – we have been hearing it so often that it is starting to become boring.
The evidence for water on celestial bodies such as Mars, Europa and Saturn’s moon Enceladus is now overwhelming. It is time to take the next step and actually look for life itself.
Full article here:
https://www.theguardian.com/commentisfree/2016/sep/28/discovery-water-jupiter-space-alien-life-europa
To quote:
When I say life, I don’t mean little green men who say: “Take me to your leader.” Microbes will do because the discovery of single alien microbe would be the greatest advance that science could ever make. We still do not know how life began on Earth. If we could find some elsewhere, we could look to see how similar or different it was from Earth life. Does it rely on DNA, for example? Do its similarities and differences hold clues about how it, and therefore we, were formed?
However, this is where the risk and the fear of failure lies. Despite what you might think from the news stories, water is no guarantee of life. Water is certainly a prerequisite on Earth and there are good scientific reasons for believing water may be essential for life of any description, but there is no guarantee that a watery celestial body will automatically play host to microbes.
Perhaps this is the reason that Nasa has seemed strangely reluctant to take the next step of actually looking for life. Earlier this century, they shied away from the Terrestrial Planet Finder, a telescope designed to look for Earth-like worlds and analyse their atmospheres for traces of life. With their Mars Curiosity rover in 2012, they specifically did not put any life-detecting equipment on board.
and…
Nasa was not always this timid. In the 1970s it sent two landers to the surface of Mars. These Viking missions carried experiments to look for life. The results were deemed inconclusive and then simply not followed up. This could have been a mistake. A recent re-analysis of the data suggested that, while the results fell short of a clear detection of Martian life, there were enough positive indicators to warrant a follow-up.
Perhaps in response, Nasa has now quietly stated that its next Mars rover in 2020 will also look for life. But at least the European Space Agency is proving bolder. In 2020 it plans to launch the ExoMars rover to Mars that will carry a suite of instruments capable of looking for the evidence of life, either past or present. That search should now be extended to Europa.
There is admirable scientific caution, and there is ducking the challenge. To go to Europa now without the ability to look for life is the latter. It is time for vision and commitment: discovering life on another planet would send shockwaves through science and fundamentally transform our understanding of our place in the universe: we must step up the search.
Will the Trump Administration get us to Europa?
http://www.dailygalaxy.com/my_weblog/2016/11/-europa-bound-trumps-space-policy-to-refocus-nasa-away-from-mars-to-explore-jupiters-moon-with-its-v.html
To quote:
The most interesting possibility of a new NASA focus on Europa is Caltech Brown’s insight that “the most interesting possibility that I can think of – is that the rocky bottom of the ocean is almost like a miniature Earth, with plate tectonics, continents, deep trenches, and active spreading centers. Think about mid-ocean ridges on Earth, with their black smokers belching scalding nutrient-rich waters into a sea floor teaming with life that is surviving on these chemicals. It doesn’t take much of an imagination to picture the same sort of rich chemical soup in Europa’s ocean leading to the evolution of some sort of life, living off of the internal energy generated inside of Europa’s core. If you’re looking for Europa’s whales – which many of my friends and I often joke that we are – this is the world you want to look for them on.”
JPL News | November 30, 2016
From Monterey Bay to Europa
If you think operating a robot in space is hard, try doing it in the ocean.
Saltwater can corrode your robot and block its radio signals.
Kelp forests can tangle it up, and you might not get it back.
Sharks will even try to take bites out of its wings.
The ocean is basically a big obstacle course of robot death. Despite this, robotic submersibles have become critical tools for ocean research. While satellites can study the ocean surface, their signals can’t penetrate the water. A better way to study what’s below is to look beneath yourself — or send a robot in your place.
That’s why a team of researchers from NASA and other institutions recently visited choppy waters in Monterey Bay, California. Their ongoing research is developing artificial intelligence for submersibles, helping them track signs of life below the waves. Doing so won’t just benefit our understanding of Earth’s marine environments; the team hopes this artificial intelligence will someday be used to explore the icy oceans believed to exist on moons like Europa. If confirmed, these oceans are thought to be some of the most likely places to host life in the outer solar system.
A fleet of six coordinated drones was used to study Monterey Bay. The fleet roved for miles seeking out changes in temperature and salinity. To plot their routes, forecasts of these ocean features were sent to the drones from shore.
The drones also sensed how the ocean actively changed around them. A major goal for the research team is to develop artificial intelligence that seamlessly integrates both kinds of data.
“Autonomous drones are important for ocean research, but today’s drones don’t make decisions on the fly,” said Steve Chien, one of the research team’s members. Chien leads the Artificial Intelligence Group at NASA’s Jet Propulsion Laboratory, Pasadena, California. “In order to study unpredictable ocean phenomena, we need to develop submersibles that can navigate and make decisions on their own, and in real-time. Doing so would help us understand our own oceans — and maybe those on other planets.”
Full article here:
http://www.jpl.nasa.gov/news/news.php?feature=6686
To quote:
Truly autonomous fleets of robots have been a holy grail in oceanography for decades,” Thompson said. “Bringing JPL’s exploration and AI experience to this problem should allow us to lay the groundwork for carrying out similar activities in more challenging regions, like Earth’s polar regions and even oceans on other planets.”
Autonomous drones could search for life in Europa’s subsurface ocean
By Laurel Kornfeld
December 3, 2016
Autonomous drones capable of making their own decisions could someday be used to search for signs of life in the subsurface oceans of icy outer Solar System worlds such as Europa. As part of an ongoing research project aimed at developing robotic submersible drones, researchers representing NASA and several other institutions recently tested six drones, using them to study changes in temperature and salinity beneath the waves of Monterey Bay, California.
Coordinated to work together as a fleet, the drones traveled for miles. They faced hazards such as potentially corrosive saltwater, entanglement in forests of kelp, and even sharks, who at times attempted to bite their wings.
In spite of these hazards, submersible drones are now frequently used for ocean research because satellites are only capable of studying ocean surfaces – their signals are unable to penetrate subsurface water. For NASA, artificially intelligent submersible drones could represent one of the best options in terms of searching for underwater signs of life on other worlds.
Full article here:
http://www.spaceflightinsider.com/space-centers/jet-propulsion-laboratory/autonomous-drones-could-search-for-life-in-europas-subsurface-ocean/
Given the signal delays plus the need to maintain a signal from below the ice, drones will need to be as autonomous as possible. Fortunately, technology seems to be making sufficient headway to make this feasible.
In the Deep, Clues to How Life Makes Light
Bioluminescent organisms have evolved dozens of times over the course of life’s history. Recent studies are narrowing in on the complicated biochemistry needed to illuminate the dark.
By Steph Yin
December 1, 2016
Dive deep enough under the surface of the ocean, and light reigns. Some 90 percent of the fish and crustaceans that dwell at depths of 100 to 1,000 meters are capable of making their own light. Flashlight fish hunt and communicate with a flashing Morse code sent by light pockets that pulse under their eyes. Tubeshoulder fish shoot luminous ink at their attackers. Hatchetfish make themselves appear invisible by generating light on their underbellies to mimic downwelling sunlight; predators prowling below look up to see only a continuous glow.
Scientists have indexed thousands of bioluminescent organisms across the tree of life, and they expect to add many more. Yet researchers have long wondered how bioluminescence came to be. Now, as explained in several recently released studies, researchers have made significant progress in understanding the origins of bioluminescence — both evolutionary and chemical. The new understanding may one day allow bioluminescence to be used as a tool in biology and medical research.
One longstanding challenge has been determining how many separate times bioluminescence arose. How many species came to the same conclusion, independent of one another?
Though some of the most familiar examples of light from living organisms are terrestrial — think of fireflies, glowworms and foxfire — the bulk of evolutionary events involving bioluminescence took place in the ocean. Bioluminescence is in fact markedly absent from all terrestrial vertebrates and flowering plants.
Full article here:
https://www.quantamagazine.org/20161201-how-life-makes-light-bioluminescence/
China’s Deep Dive! Unmanned Submersibles Reach the Hadal Zone –“Deeper Than Mount Everest, Preview of Jupiter’s Ocean-Moon Europa”
December 29, 2016
In 2011, international team of microbiologists of the Microbiology of Extreme Environments Laboratory in partnership with the Institute of Oceanography of Xiamen (China) and the Earth Science Laboratory discovered a new species of archaebacteria, Pyrococcus CH1,discovered thriving on a mid-Atlantic ridge within a temperature range of 80 to 105°C and able to divide itself up to a hydrostatic pressure of 120 Mpa (1000 times higher than the atmospheric pressure).
The piezophilic microorganisms constitute a subgroup of extremophiles. Discovered on the site “Ashadze”(2) at 4100 meters depth, the deepest vent field explored so far, the CH1 strain was successfully isolated and assigned to the genus Pyrococcus, within the Euryarchaeota lineage of the Archae domain. The discovery extended the known physical and chemical limits of life on Earth.
Full article here:
http://www.dailygalaxy.com/my_weblog/2016/12/chinas-deep-dive-unmanned-submersibles-reach-the-hadal-zone-deeper-than-mt-everest-preview-of-a-prob.html
To quote:
Extraterrestrial life is the most interesting thing ever, bar nothing. We have two possible life-locations right here on our solar system doorstep – but we have to choose which to check. We want to go everywhere, but with a price-tag of billions of dollars per outer-planet probe we have to decide and flipping a coin just won’t cut it.
Option Number One is Jupiter’s Europa, the favored satellite son of many exobiologists and even Arthur C. Clarke himself. While distinctly non-Terran, huge sub-surface lakes probably heated by tidal stresses, and even an extremely tenuous oxygen atmosphere make it a leading contender.
This British scientist says we will find life on either Europa or Enceladus or maybe both in 2017 due to Donald Trump. Hey, stranger things have happened… I think:
https://www.inverse.com/article/25867-aliens-cassini-europa-enceladus-simon-foster
To quote:
At any rate, after making these points Foster proceeds to tell the Daily Mail that Trump might actually be good for science and alien-hunting because it will simply be the practical investment to make, so you’d be forgiven for not getting quite as hyped about this latest round of aliens-or-maybe-not-but-definitely-stay-tuned.
“I think when he gets in he will be different,” said Foster. “Because from a business point of view, he is going to look at things like hurricanes and how they’ll affect America, so you are going to get more intense and frequent and people are losing their homes and businesses are being ruined by this, he is a businessman and that will cost you tax I think he will take a more pragmatic view on this.”
Well this is certainly going to be useful technology for an Europan submersible probe:
http://www.whoi.edu/oceanus/feature/communicating-under-sea-ice
JPL NEWS | FEBRUARY 8, 2017
NASA Receives Science Report on Europa Lander Concept
A report on the potential science value of a lander on the surface of Jupiter’s icy moon Europa has been delivered to NASA, and the agency is now engaging the broader science community to open a discussion about its findings.
In early 2016, in response to a congressional directive, NASA’s Planetary Science Division began a pre-Phase A study to assess the science value and engineering design of a future Europa lander mission. NASA routinely conducts such studies — known as Science Definition Team (SDT) reports — long before the beginning of any mission to gain an understanding of the challenges, feasibility and science value of the potential mission.
In June 2016, NASA convened a 21-member team of scientists for the SDT. Since then, the team has deliberated to define a workable and worthy set of science objectives and measurements for the mission concept, submitting a report to NASA on Feb. 7.
The report lists three science goals for the mission. The primary goal is to search for evidence of life on Europa. The other goals are to assess the habitability of Europa by directly analyzing material from the surface, and to characterize the surface and subsurface to support future robotic exploration of Europa and its ocean. The report also describes some of the notional instruments that could be expected to perform measurements in support of these goals.
Full article here:
http://www.jpl.nasa.gov/news/news.php?feature=6737
To quote:
The concept lander is separate from the solar-powered Europa multiple flyby mission, now in development for launch in the early 2020s. The spacecraft will arrive at Jupiter after a multi-year journey, orbiting the gas giant every two weeks for a series of 45 close flybys of Europa. The multiple flyby mission will investigate Europa’s habitability by mapping its composition, determining the characteristics of the ocean and ice shell, and increasing our understanding of its geology. The mission also will lay the foundation for a future landing by performing detailed reconnaissance using its powerful cameras.
The report is online here:
http://solarsystem.nasa.gov/docs/Europa_Lander_SDT_Report_2016.pdf