Mention Robert Zubrin’s name and the planet Mars invariably comes up, given his long-time work on finding ways to establish a human presence there. Dr. Zubrin is the originator of Mars Direct, the author of The Case for Mars, and founder of the Mars Society. But his work on interstellar matters is likewise significant, including the analysis, with Dana Andrews, of the Bussard ramjet, which taught us much about magsails and drag, offering a useful way to re-think starship braking at destination. Another key Zubrin creation is the nuclear salt water rocket concept. With over 200 papers and five books to his credit, he runs Pioneer Astronautics, where he continues to focus on innovative aerospace technologies. Today’s essay goes in another direction, with a fresh look at interstellar communications using microscopic data carriers. Ponder now how information can be conveyed star to star, and how we might find it by methods far removed from conventional SETI.
by Robert Zubrin
Abstract

Since the dawn of the SETI effort in 1960, it has been generally assumed that the transmission of information across interstellar distances can most practically be accomplished using electromagnetic waves, with the most popular candidate method being radio in the 21-cm wavelength (1.42 GHz) range. Accordingly, a series of searches based on this assumption have been conducted, thus far without any success. In this paper, we will advance a hypothesis that the reason for this failure is because radio is, in fact, a very inefficient means of interstellar communication between species, and that a superior alternative is available. Specifically, we will show that communication between species can be much more effectively accomplished over interstellar distances using microscopic high density data storage packages sized between 1 and 10 microns. Such packages have already been detected. They are, in fact all around us, and within us, in vast numbers and varieties. Generally known as bacteria, these spaceflight-capable data storage systems are carrying enormous amounts of information, only a small fraction of which has any identifiable purpose. Could interstellar messages be found encoded within the genomes of microbes? Could records of such past transmission be found within the genomes of multicellular organisms? In this paper we shall explore the possibilities, discuss how such transmissions could be efficiently sent, and propose methods by which such a hypothesis might be falsified or verified.
Introduction
It is a general principle of science that the laws of nature should apply equally well throughout the universe. Specifically, since the dawn of modern science in the Renaissance under the philosophical banner “as above, so below” it has been taken as axiomatic that the laws of nature that prevail elsewhere in the universe should also apply on Earth, and those that apply on Earth should apply equally well elsewhere. This being the case, it necessarily follows that if life and intelligence could develop from physics and chemistry via natural processes on Earth, it should also have done so in innumerable other equally satisfactory physical and chemical environments throughout the cosmos. Indeed, the early Earth at the time of life’s first appearance immediately following the end of the heavy bombardment was in no way exceptional. Furthermore, the processes by which life can develop from simple forms to complexity and intelligence are, in broad outline, well understood. While the universe is vast in space, it also is so in time, so that a spacecraft traveling at a velocity of 0.0001 c (the speed of the Earth in its travel around the Sun) could in 4.5 million years (0.1% the age of the Earth) travel 450 light years, a radius encompassing approximately 1 million stars, with surely enough candidates for many additional origins for life and civilization. Despite these favorable odds, no such extraterrestrials have been detected, a mystery leading the physicist Enrico Fermi to ask his famous paradoxical question at a 1950 Los Alamos lunchtime meeting: “So then, where are they?”
One possible reply to the Fermi Paradox is that they are out there, but that if you want to find them you need to look for them in the right way. In 1959, Cornell physicists Phil Morrison and Giuseppe Cocconi (Cocconi and Morrison, 1959) proposed that extraterrestrials might be communicating across space using 1.42 GHz radio, as the emissions of hydrogen gas at that frequency make it the most listened-to band in radio astronomy. Moreover, it is approximately the same frequency as the L-band and S-band radio systems that were becoming state of the art for spacecraft communication at that time, a fact which added credence to the Morrison-Cocconi hypothesis, and made it readily testable as well. Accordingly, shortly thereafter astronomer Frank Drake attempted to detect such signals from Tau Ceti, without success. Undeterred, Drake, his co-workers, competitors, and successors continued with many such search searches, from that time down to the present, where the SETI Institute, among others, is continuing the effort on a greatly expanded basis with vastly improved instrumentation, but no better results.
Given this failure, it is appropriate to revisit the assumptions behind the Morrison-Cocconi hypothesis suggesting interstellar communication via S-band. Certain of its supports have already been falsified by technological progress, in that, a mere 60 years later, S-band is already obsolete. Instead our spacecraft now communicate at higher frequencies, such as X-band (10 GHz) and Ka-band (30 GHz), as these higher frequencies allow for higher bandwidths for spacecraft communication systems of a given size and power.
But while the Morrison-Cocconi hypothesis, and resulting search, can be adjusted to take into account such improvements, there are deeper problems. The first of these is that communicating effectively across interstellar distances via radio is incredibly hard and inefficient. To see this, let us consider what it would take to design such a system.
The Mars Reconnaissance Orbiter (MRO), launched in 2005, has a modern 100 W X-band communication system. Equipped with a 2-m diameter dish, it can transmit at a rate of 6 Mb/s to a 70-m receiving system on Earth at a distance of 100 million km. A modest range for effective interstellar communication would be 10 light years, or 100 trillion km. At this million-fold greater distance, the MRO communication system would have its data rate reduced by a factor of a trillion, from 6 Mb/s to 6 microbits/s, or 200 b/year. This would appear to be too slow for practical purposes, so let’s upgrade the transmitter power to 1 GW and its dish size to 70 m. Taken together these upgrades would increase the data rate by a factor of 10 billion, to 60 kb/s, which would be fine. The upgraded system would have a capability of 600 b/s at 100 ly, which is still sufficient to be useful, as those who can remember working with computer modems in the 1980s can readily attest.
Assuming that the transmitter was X-band, the diameter of the beam at 10 ly would be about 2500 AU, so it would encompass the whole solar system, and then some, but the closest neighbor solar system would be 100 times further away that the width of the beam. So ET’s 1 GW transmitter could only be used to signal one solar system at a time. How would they know who to signal?
A good place to start would be to only send signals to planets manifesting a biosphere. These could be detected using astronomical techniques by observing the spectral signal of free oxygen in the planet’s atmosphere. But Earth, for example, would have provided a positive answer to such a search criterion for the past 500 million years, but only has only possessed a species able to receive and detect such a signal for the past 50 years. Based on these odds, ET would have to set up 1 million transmitters to living planets have a 10% chance that one of its signal units would be sending at the right time. This would require a transmitter system power of 1000 TW, about two orders of magnitudes higher than the total power produced by human civilization today. If they wanted to keep their odds as good, but reduce the number of transmitters, they could do so, but only at the cost of continuing to power the transmission program for millions of years. Furthermore, to receive the signal, the inhabitants of the target planet would have to be focusing their 70-m dish at precisely the right star at the right time, be listening at the right frequency, and be technologically and intellectually prepared to recognize and decipher the signal. Using lasers instead of radio to transmit would reduce the power requirement significantly, but it would still be huge, and furthermore, it would impose a requirement that the receiving civilization have a giant telescope focused on the transmitter’s narrow beam, at the right time, with its optics limited to the transmitting frequency to avoid having the transmitter outshined by its home system’s star. This seems like a rather farfetched hope upon which to expect ET to spend such a large investment in infrastructure and energy.
Surely there must be a more efficient approach. What could it be?
Interstellar Data Transmission by Microbial Storage Drives
One problem of data transmission by radio is that it occurs in real time, leaving no record behind. Once the transmission is over, it’s gone. If the intended recipients miss it the first time, they’ve missed it for all time. As a result, the transmitting party is forced to transmit repeatedly, perhaps endlessly, in the hope that one at least one occasion, someone might be listening.
Imagine that you want to tell a story to children. So you walk onto the front porch of a house and tell the story, whether any children are there are not. In fact, they are usually out running around, so you probably have missed your audience. But the odds against you are worse, because while the house is suitable for children, they may not have been born yet, or those that are there may be too young to understand your story, or they could have grown up and moved out. Now, you could increase your odds by going from porch to porch, reading the story again and again in the hope that someone might be there to listen. But this could get exhausting. A better strategy would be to go about slipping story books into houses through their mail slots. Even if most of them ended up on the shelf or sold to used book stores, sooner or later many of them would likely get read. The only problem with this strategy is its cost; giving out a lot of storybooks could be expensive. But if you could get them for free, and have them delivered for you cheaply, it could be a very good approach, allowing you to reach many children not only today, but for decades and generations to come.
In short, I am suggesting, in agreement with Davies (Davies, 2010), that rather than transmit information across interstellar distances using radio waves, that solid objects containing records be used instead. This may seem like it would be very inefficient, but in fact every planetary civilization orbiting a star has available to it an engine that it can use to send information across space at little or no energy cost. This is the star itself.
Let us consider our own Sun as a case in point. At 1 AU, the sun attracts objects to it with a gravitational acceleration of 0.006 m/s2. It also repels objects from it via its light pressure with a force of 0.000009 N/m2. If these two forces are equal, and object will feel no attraction from the Sun, and fly out of the solar system in a straight line with the Earth’s velocity of 30 km/s. (Both of these forces change with the inverse square of their distance from the Sun, so if they are equal at 1 AU, they remain equal at any distance.) Assuming the object has a radius r and a density d, and setting these two forces as being equal we find:
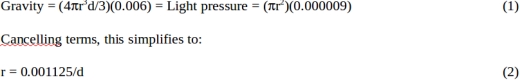
If d = 1000 kg/m3 (water density), we find that r = 1 micron. This means that such an object would have a diameter of 2 microns. However, it is not necessary to cancel all the force of gravity to escape the solar system. If half of the gravitational attraction is cancelled, an object with the orbital velocity for the full gravity object will escape. In the case above, this would imply a maximum diameter of 4 microns.
If the star were brighter than the Sun compared to its gravity, which would be the case with F stars, the objects could be somewhat larger. If it were dimmer than the Sun compared to its gravity, as would be the case with K stars, the objects would be need to be smaller. If their initial orbits were elliptical, rather than circular, the objects could be bigger. So, depending on assumed conditions, the diameter of the objects could range from 1 to 10 microns, and be readily projectible across interstellar distances using no other mechanism than the pressure of the star’s light. This is precisely the size range of many typical bacteria. (Arrhenius, 1908) It is also possible that the solar wind, which moves at a velocity of about 500 km/s, could be used to propel particles out of the solar system at very high velocity. However, in order for a particle to interact effectively with the solar wind it would need be strongly magnetized, allowing it to function as a miniature magnetic sail. (Zubrin and Andrews, 1989.) Under normal circumstances even such a strongly magnetized particle would be propelled away from the Sun by the solar wind with less force than that provided by light pressure. During solar flare events, which greatly increase the force of the plasma wind emanating from the Sun, however, this could change radically.
In Table 1, we show the relationship between star type, population fraction, star mass, luminosity, orbital distance and velocity and maximum particle diameter for stellar system escape for stars of various types. In each case the launch planet is assumed to be in a circular orbit in the given star type’s habitable zone. It can be seen that for type F, G, and K stars, collectively amounting to about 22.5% of the stellar population, that starlight can readily propel bacteria sized objects to system escape velocity. At half the diameters shown the objects will be projected outward from the stellar system at the orbital velocity given, at less than half, still greater velocity. Objects larger than that shown will not escape the system by light pressure alone, but could still do so if the light pressure is enough to drive them into an elliptical orbit that intersects a large planet capable of delivering a gravity assist.
Upon reaching a destination solar system, the bacteria-sized particles could be decelerated using the same mechanism.
Table 1 Interstellar Particle Transmission Capabilities of Star Types.
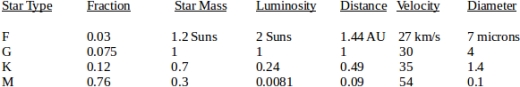
In Table 1, the particles are assumed to be simple spheres. If more complex shapes are used, for example shapes with a spherical core surrounded by thinner wings, the core spheres can be thicker and still achieve the same velocities. For example, a sphere with a diameter of 4 microns surrounded by a disc with a diameter of 8 microns and a thickness of 1 micron would have the same surface/mass ratio as a simple sphere with a diameter of 1.2 microns, and would therefore be able to escape a K star. Such “microsailcraft” designs could be readily mass-produced artificially or potentially created through natural crystallization processes, as exemplified by snowflakes. They could be stabilized in a manner to effectively serve as sails either by spinning (as some snowflakes do), or by having inherently stable shapes, as exemplified by a badminton birdie.
So, bacteria can be projected across interstellar space at essentially no power cost to the transmitting party, beyond that required to launch them to planetary escape velocity. The latter could be accomplished either by artificial technological means that are well within our means today – and therefore clearly feasible for advanced extraterrestrials – or potentially through natural processes such as asteroidal impacts. They also may be cheaply mass produced. (McDowell, 2003) But can they carry useful information?
The answer is most certainly yes. The genetic material of individual common bacteria is estimated to contain between 130 kB to 14 MB of information. Current estimates that bacteria can be used to store data with a density of 900 Terabytes per gram, about 500 times the current state of the art electromagnetic hardware. This means that a bacterium 5 microns on a size could store about 120 kB of information. (Wilkins 2010, Herkewitz 2016) Taking 60 kB as typical, this means that a single bacterium can carry a record of information about equal in size to a 10,000-word (~30 page) booklet. In experiments done to date, scientists have demonstrated such capabilities by encoding entire books in DNA, and showing that bacteria can be made to replicate encoded information when they reproduce.(Ayre 2012). Most recently, researchers at Columbia University and the New York Genome project have shown that they can encode information with a density 215,000 Terabytes of information per gram in DNA, with some of the items successfully encoded including a movie, a computer operating system, and an Amazon gift card. (Service, 2017)
Traveling at a velocity of 30 km/s (0.0001 c), bacteria would take 100,000 years to fly 10 light years. This would expose them to cosmic ray doses between 1 and 10 Mrads, which is close to the limit for survivability of hardy microcrobial species such as radiodurans. This need not be a show stopper. A message sent using bacteria storage would no doubt use billions of individuals, and if even a few survived the trip the message could still get through. While ultraviolet light would kill unshielded bacteria in days, effective shielding against this hazard can be provided by a half micron of soot. (Hoyle, 1981). Such protection would be provided by design in any artificial microsailcraft, but could conceivably also occur naturally.
Such long trips might not be necessary, however. Once they reach a planet, bacteria will multiply to vast numbers. They can then be ejected again into space via cometary impact. Such impacts are most likely to occur during periods when a foreign star is passing through the Oort cloud of the bacteria’s home star, as such a passage would destabilize Oort cloud object orbits orbiting both stars, and thereby causing impacts to occur. Like frigates in the age of fighting sail, which could span the globe with their movements but only reach a few hundred yards with their guns, roving solar systems discharge their broadsides at each other only during rare close approaches. As a result, ejected bacteria might typically only need to travel distances on the order of 0.1 ly to reach a new planetary home, with radiation doses accordingly reduced by two orders of magnitude compared to those postulated above. Given the density of stars in our own region of the galaxy, and assuming a random velocity of stars relative to each other of 5 km/s, it can be shown that a star is likely to experience such a close encounter about once every 20 million years, a frequency strikingly close to the observed mean time of 26 million years between of mass extinctions on Earth. (Zubrin, 2001). It may be noted that microbes traveling embedded in impact debris would be well shielded from ultraviolet and soft x-rays, thereby increasing their survival odds. (Melosh, 1988, Hornek, Klaus, and Mancinelli, 2010)
The Milky Way galaxy is 13 billion years old. Allowing 3 billion years for several generations of early stars to seed the place with heavy elements, that leaves 10 billion years, or 500 stellar close-encounter doubling times of 20 million years each for life to spread from its first planet of origin to everywhere else. So the answer to Fermi’s paradoxical question is almost certainly this: They’re here.
The Purpose of Interstellar Communication
At this point, we need to reexamine the question of what might be the purpose of interstellar communication. Certainly, if a species were spacefaring and had sent out expeditions which established settlements in nearby solar systems, it would want to maintain communication to exchange or trade information between its various worlds. For such purposes, high-gain directed electromagnetic transmissions would be the most practical, as they are the fastest possible, the most secure, and all the required technological and linguistic conventions would be known and mutually understood between the parties involved.
But if we are talking about communication between different species originating in different and distant worlds, what is the point? In speculative SETI literature, it is frequently supposed that there are intelligent aliens out there, who for some reason want others to know that they exist, and therefore transmit signals such as the value of into the void, so that other smart folks won’t confuse them with astrophysical phenomenon. Then, assuming that someone picks up the signal, they will transmit back the value of e, or the golden mean, or some other special number, thereby completing the freemasonic handshake. This done, the two parties could then proceed to methodically expand their mutual vocabulary, eventually allowing them to exchange QST cards, recipes, celebrity gossip, novels, scientific theories, starship designs, and treaties of alliance against the barbarians from the Galactic rim.
However, as noted above, both experimental searches and theoretical considerations suggest that such a picture does not correspond to reality. Microbial data transmission is possible, but it does not lend itself to conversations of the types described above. Rather it is a superior method of interstellar broadcasting.
So the question is, what kind of information is really worth broadcasting – that is distributed as widely as possible to people who we don’t know and are not likely to hear back from? If human experience is any guide, the answer is propaganda. Think of Radio Free Europe, or its Cold War opposite number, Radio Moscow, for example, and their persistent messaging: “We are good. You should admire us. You should be like us. You should join us.” Another example would be the Gideons, placing Bibles in hotel rooms in the hope that their unknown readers would see the light and become Christians. We also try to broadcast ourselves to worlds we will never see, by using art to send our message across deep time. Thus Pericles at the Parthenon: “Future ages will wonder at us, even as the present age wonders at us now.”
The key to propaganda is in the root of the word itself: propagate. Through propaganda we seek to propagate ourselves across both space and time. This can be in spirit, as in the cases described above, or in the flesh, through physical reproduction. Indeed, while only a relative handful of people have been able to message the future through monuments, literary works, or art, the great majority of those of the past world who have sent us something of themselves have done so by propagating themselves biologically. Using this method, they have transmitted to our world not only their genotypes and phenotypes, but even their languages, beliefs, and traditions as well. Propagation is propaganda. Propaganda is propagation. It is the most desired form of communication. It is how the past has communicated with us, and how we seek to communicate with the future. This is a key point, because interstellar communication through any means must perforce be communication across time.
So, it should be clear. If we are going to transmit across the ages, we need to send instructions on how to make ourselves. Such messages are not sent via radio. They are sent using genes.
The code of life is the code of the cosmos.
Panspermia or Geospermia?
It is a striking fact that, despite several centuries of microbial research by thousands of competent investigators, no free-living organisms have been found on Earth that are simpler than bacteria. This is truly remarkable because, as simple as bacteria may be compared to more complex organisms, they are certainly not simple in any absolute sense, incorporating as they do, among other things, the entire elegant double-helix scripted language of DNA. Believing that bacteria were the first life forms to emerge from chemistry is like believing that the iPhone was the first human-invented machine. This is incredible. Just as the development of the iPhone had to be preceded by the development of computers, radio, telephones, electricity, glassware, metallurgy, and written and spoken language, to name just a few necessary technological predecessors, so the creation of the first bacterium had to be preceded by the evolution of a raft of preceding biological technologies. But we see no evidence of any such history. We still see devices all around us that use one or more of the iPhone’s ancestor technologies – telephones, light bulbs, batteries, glass windows, and steel knives, for example – but we see no pre-bacteria organisms. This observation has led many investigators, dating back to Arrhenius (Arrhenius 1908) over a century ago, to postulate that life on Earth is an immigrant phenomenon. According to this “panspermia” hypothesis, bacteria did not originate on Earth, but came here from space, after which they gave rise via generally understood evolutionary processes to all other life forms.
The panspermia hypothesis is generally disliked by origin of life researchers, because it completely ducks their central question of how life originated from chemistry in the first place. This is particularly the case for the original form of the panspermia hypothesis offered by Arrhenius, who believed that the universe and life had existed eternally, thus making the question of the origin of either meaningless. However, if the panspermia hypothesis is taken to simply open the question as to the location of life’s planet of origin, then it is by no means useless. Consider: an investigator seeking to explain the origin of Americans would be crippled in his or her research if he or she had to accept as axiomatic the conceit that humans evolved independently in North America (and even more so if Golden, CO were specified.). No, the fact is that humans originated in Africa, and only came to the Americas much later. This is why we can find evidence of humanity’s closest relatives, primate ancestors, and earliest cultures and technologies in Africa but not in North America. Knowing this, an investigator would not be bound to try to explain the independent origin of humans from native North American (or Goldenian) fauna, but instead could focus on conditions and biological foundations that were present in Africa in the relevant period. Similarly, there have been origin of life experiments, such as the famed Miller-Urey experiment, that have been discounted because they postulated conditions that did not exist on the early Earth. If the possibility of an extraterrestrial origin of life is accepted, such objections lose their force.
Indeed, insistence on geospermia by assumption puts origin of life researchers in the same absurd position as the above described unfortunate paleontologist, whose assumption of a local origin for humanity forces him or her to reject the theory that humans evolved from higher primates because there were no such species in evidence in Golden, CO at the time of humanity’s appearance. There are innumerable planets where the spontaneous formation of amino acids from chemistry, as demonstrated by the Miller-Urey experiment, could readily have occurred, as opposed to the early Earth, where it could not. Science needs to follow the data, not defy it. Therefore it is the Miller-Urey experimental results that discredit the assumption of geospermia, rather than the reverse.
Further support is offered to the panspermia hypothesis by discoveries of bacterial fossils known as stromatolites, dating back approximately 3.5 billion years, and residues of biological activities dating back 3.8 billion years, that is practically right back the end of the heavy bombardment that previously made the early Earth uninhabitable. In fact, as this is being written, a team of researchers have just reported microfossils that date back 4.28 billion years, that is to the middle of the heavy bombardment. (Drake 2017) In short, life appeared on our planet virtually as soon as it possibly could (and possibly several times, before it could last), suggesting that it was already around, waiting to land and spread as soon as conditions on the ground were acceptable.
The primary counter argument offered against the panspermia hypothesis is that there may once have been prebacteria on Earth, but that they have since been wiped out by their more developed descendants. While this may be possible, it is not consistent with the history of life on Earth, in which simpler forms generally continue to exist in abundance even after they give rise to higher or more complex varieties. In any case, this argument is only an excuse for the lack of any evidence for any prebacterial history of life on Earth. Accordingly, it has no power or potential to falsify the panspermia hypothesis.
Furthermore, it needs to be understood that the conceit that life originated on Earth is quite extraordinary. There are over 400 billion of stars in our galaxy, with multiple planets orbiting many of them. There are 51 billion hectares on Earth. The probability that life first originated on Earth, rather than another world, is thus comparable to the probability that the first human on our planet was born on any particular 0.1 hectare lot chosen at random, for example my backyard. It really requires evidence, not merely an excuse for lack of evidence, to be supported.
The panspermia hypothesis could be falsified however, if we were to send explorers to Mars and find either a) no evidence of any past or present life, b) evidence for the presence of prebacteria or c) evidence of life of sufficiently different type as to imply a second genesis. Condition (a) would falsify panspermia because Mars had liquid water on its surface during the period when life appeared on Earth, so that if Earth were seeded via panspermia, Mars should have been seeded too. Condition (b) would refute panspermia directly by revealing the prior evolutionary history of Earth life on Mars, from whence it could readily have been transmitted here via meteoric impact. Condition (c) would refute panspermia by showing two independent origins. However, if none of these conditions are met, and we find evidence of past or present bacteria on Mars similar in structure to Earth bacteria dating back to the planet’s early history, with no evidence of prebacteria, the panspermia hypothesis would be strongly supported.
In the absence of falsification, we are presented with three possibilities for interstellar microbial transmission.
1. The transmission is natural, being the result of ejection of material from microbe-inhabited planets following meteoric impacts.
2. The transmission is artificial, being the result of intentional dispersal by intelligent extraterrestrials of microbes carrying imprinted encoded information.
3. The dispersal is both artificial and natural, being the result of both processes listed above going on simultaneously.
With respect to the above listed possibilities, the one that seems most difficult to defend is (2), because if bacteria can survive interstellar trips, there will be natural transmission, regardless of whether artificial transmission is also going on. Indeed, it is hard to escape the conclusion that natural transmission has been going on for at least 3.6 billion years, if from no other original source than the Earth. If the average time between close stellar encounters is 20 million years, with number of microbe inhabited system doubling each time, we could expect 2180 solar systems in our galaxy to have been Earth-progeny-microbe-invaded by now, which is to say all of them, many times over. (This being the case, the probability that the Earth was actually the first of these billions of microbe-inhabited worlds would be vanishingly small.)
So, the question is not whether interstellar microbial transmission is going on; it almost certainly is. Further, even if is not occurring naturally, it is still clearly possible through artificial means. So indeed, far from being a necessary condition for microbial SETI, even the possibility of natural panspermia creates difficulties, as it introduces the potential for noise that could drown out the signal.
The key question for microbial SETI is whether there is artificial intelligent input being inserted into the vast flood of genetic information traveling around the Earth. Are there any real letters of importance to be found in the deluge of junk mail? If so, how could we pick them out?
Distinguishing ET’s Messages from Junk Mail
Radio SETI researchers record electromagnetic waves received from space, and examine them with algorithms to try to detect something that seems too organized to be merely the result of astrophysical phenomenon. In principle, microbial SETI researcher could take a similar approach, using gene sequencing technology to process large numbers of bacterial genomes, looking for something that just doesn’t seem natural. Perhaps buried amidst the junk DNA of some bacterial species is a sequence of amino acids that an expert cryptanalyst can decode to read; “Here is the value of π, 3.14159265…This is just our way of saying hello. You will find the primer for our language on the next gene, and starship design instructions on the gene after that.” Perhaps that is a somewhat facetious way of putting it, but certainly if we believe that extraterrestrials want to send us some such signals, a direct attempt to sort through the mail pile using gene-sequencing and cryptanalytical tools to see if anything interesting pops up might be warranted.
That said, there is, as noted above, grounds for skepticism that this is the form of communication that intelligent species would find interesting. If there is a field of life throughout the galaxy, initiated on innumerable worlds by natural panspermia, it could be expected to evolve in multitudes of new and unpredictable directions through natural processes including mutation and natural selection, driven by chance and diverse environmental conditions. It seems to me that the most portentous form of communication that intelligent extraterrestrials could undertake would be to try to propagate themselves by sending out genetic information to influence this chaotic process in their own direction. Genomes can contain dormant plans for complex traits, as evidenced by recent work in which scientists activated what had been considered junk DNA in chickens to produce long-lost dinosaur features, like teeth. (Hoggenboom 2015, Bhuler 2015) Well, chickens are descended from dinosaurs, so perhaps it’s not too astonishing to discover that they still keep some of the old body plans on file. Such plans could come in handy if new conditions require a radical evolutionary leap. But could it be possible that some such genetic plans were sent here intentionally by bacterial conveyance? Could some have been used, and others be still awaiting their chance? There are a variety of physiological features, such as the complex eye, or bird wings, whose origin is hard to explain in terms of incremental natural selection, as they appear to be completely non-functional in partially-developed form. The raising of such paradoxes has long been a stock in trade of theists arguing the case for supernatural “intelligent design.” But while there are, by definition, no supernatural phenomenon, there really are many things – including not only buildings and ships, but also domesticated animals and plants – that are the product of intelligent design. Bacteria can transfer genes among themselves, and to and from macrofauna and macroflora (Yong, 2016, Margulis and Sagan, 2008). Instead of sending us greetings and saluting us with the value of π, could extraterrestrials be sending us microbial messages for the purpose of guiding the evolution of our biosphere?
As fantastical as it sounds, I believe this is a testable hypothesis. Specifically, one could search for genetic material being carried by bacteria that can be inserted into animals or plants and result in the production of striking adaptations that have not yet manifested themselves in living species on Earth. Birds once had teeth, but they never had radar. No species is known which communicates telepathically using bioradio. There are any number of useful adaptations that are physically possible but which never have manifested themselves in terrestrial biology. Furthermore, there are traits that we do see in some species, but not in their ancestors. Mice were once fish, but fish were never mice. The fact that mice still carry fish traits has been clear since the 19th Century, when it was observed that mammal embryos exhibit fish traits such as gills. But do fish carry in their inactive DNA advance plans for mammalian traits? It is possible that such traits could be induced by the transference of bacterial genes. But from what source? If such were found, could they have been sent by extraterrestrials in the distant past, to either fish or their ancestors? Or did fish get them from mammals by natural local bacterial transference much more recently? How could these two possibilities be sorted out?
We could also consider the past. There were certain periods when a massive amount of evolutionary innovation occurred within an extremely brief period of time, with the Cambrian Explosion and the early Eocene immediately following the KT extinction immediately coming to mind. Can the sudden appearance of so many radically new traits be best explained by the prior existence of readily available genetic plans? Or perhaps could it be the result of the opening of expanded channels of communication between genetic plans? (Hoyle, 1981)
Current evolutionary neo-Darwinian “fundamental dogma” holds that all traits of a species are only passed on genetically, so that traits acquired in life are not inherited. Furthermore, new genetic traits are only created through mutation and passed down within a species own line of descent, and that only if they pass the test of natural selection within that species line. While useful in understanding biological evolution, this theory is clearly false when applied to human social evolution. This is so because in contrast to what seems to be the case with other animal species, humans can inherit acquired traits, such as technologies, and furthermore, can inherit such traits from unrelated persons or groups. As a result, such theories as national social Darwinism, (Bernhardi, 1912, Hitler, 1941) which postulate history as a battle of nations for limited resources, with advances occurring via the resulting elimination of the less militarily proficient, are not only morally reprehensible, but scientifically wrong, since inventions made in any nation can (and generally do) ultimately benefit all nations. Indeed, were it the case that technological innovations were not laterally transferable from person to person, tribe to tribe, and nation to nation, it is doubtful that humanity would ever have advanced beyond the old stone age.
Conversely, it is easy to see the quantum acceleration of human progress following the establishment of global communications through the development of the printing press and long-distance sailing ships circa 1500, and their further improvement via railroads, steamboats, telegraphs, telephones, radio, TV and the internet in the period since. Furthermore, it is unquestionable that the growth of human population has contributed to this trend, since the more people there are, the more inventors there can be, and inventions are cumulative. This is why, contrary to Malthus, as the world’s population has gone up, the standard of living has gone up, not down. For similar reasons, it is in the self-interest of intelligent species to promote the creation of and communication with as many intelligent species elsewhere as possible. The more sources of transferable invention that there are, the more inventions each will receive, and the greater will be the power of each to add further inventions in turn. Furthermore, most intelligent species must be aware that their self interest lies in increasing, rather than decreasing, the number and effectiveness of the creative agents that they draw upon, because if they were not, they could not survive for long.
This said, can it really be true that the biosphere is so defective that it is incapable of allowing analogous transfer of useful genes between species? The creation of new useful traits by random mutation is a slow process. Clearly it would benefit all species, and thus the biosphere as a whole, were each to be able to draw to some degree on the genetic innovations of the rest. A community of life that had such ability would have greatly improved odds of survival, and enjoy tremendous adaptive advantage over any that did not. In fact, contrary to the neo-Darwinian fundamental dogma, the biosphere has exactly such a capability. Such transfers from species to species are enabled by bacteria, (Hotopp 2011). Such bacterial transfers of entire genes allow the evolution of valuable new inheritable traits to appear in species not over millennia, but within the lifetime of a single individual. Yong (2016) reports many such instances, for example species of woodrats that upon acquiring certain bacteria, immediately gain the ability to digest various plants, such as creosote or cactus, that they did not have before, thereby becoming able to live and prosper in environments dominated by such species. These valuable bacterial-derived traits are then passed from mother to child during birth, along with samples of the maternal bacteria. But since the rats are constantly broadcasting their bacteria through exhalation, these traits can be transferred to other unrelated individuals and even other species. Other bacteria have been shown to provide various species with inheritable defenses against parasites such as nematodes. Margulis and Sagan (2008) report many additional examples, including cases where the acquired genes eventually move from the bacterial microbiome carried by the animal into the animal cells themselves.
To use an analogy, the genotype of an animal’s cells is its hardware, but an animal’s characteristics are also determined by its bacterial software, or microbiome, which can be rapidly changed in and out. Evolution occurs not by random changes in the hardware circuitry, but by adding or subtracting software programs, which are constantly being developed and exchanged in vast amounts by the extremely prolific bacteria. If found by natural selection to be valuable over the long haul, these programs can end up being written into the hardware.
If the software is found to be useful, the animal’s chances of survival are improved, and the trait is passed on to her descendants. But not only that, she and each of her progeny become agents for spreading the useful trait throughout the biosphere through their exhalations and excretions. As you read this article, dear reader, you are exhaling half a million of your bacteria every minute, providing every creature around you with samples of what you and your ancestors have found to be useful. Your dog or cat is broadcasting similar service. Thus the web of life on our planet shares its inventions.
Can bacteria perform the same role from world to world? If they could, they would be a tremendously positive influence on evolution throughout the galaxy, transferring entire encyclopedias of hard won biological knowledge between planetary biospheres. But while benefiting the progress of life, it is hard to see how such capabilities benefit the bacteria themselves. Could they be serving the purposes of others? Perhaps they are, particularly since it is easy to see how others could be readily sending them.
It may be objected that ET’s wishing to control evolution elsewhere in order to reproduce themselves on other planets could hardly accomplish such as objective by spreading microbes carrying their genes, as evolutionary processes occurring on destination worlds would certainly carry matters forward in unpredictable directions. This is unquestionably true. However, returning to our example of trying to talk to the children in the neighborhood by leaving books around, we can imagine a more general method of communication. Let’s say the book is Harry Potter and the Sorcerer’s Stone. There are several layers of messaging in the book. These include:
1. Being a wizard beats being a muggle. So try to get into Hogwarts, and be sure to choose a Nimbus 2000 for your broomstick.
2. Virtue will be rewarded, vice will be punished, and good will triumph over evil in the end.
3. Reading is fun!
As a (1) method for transforming children into wizards, Harry Potter fails. It can contribute to (2) moral instruction, however, and succeeds brilliantly as (3) a way to get children to become readers. After that, there is no limit to what they might learn or become.
By analogy, the purpose of microbial data transmission might not be to direct evolution in particular ways, but simply to encourage evolution to be more creative.
Finding the Most Likely Suspects
Given these observations, how can we make use of them to detect ET? A good place to start might be to focus study on those bacteria which show the strongest signs of most recent extraterrestrial origin. These would be those best adapted for spaceflight. In nature, adaptations for particular purposes always come at a cost, and therefore generally disappear over time if they become unnecessary. Some bacteria, such as radiodurans, have excellent astronautical adaptations, including extreme resistance to hard radiation and vacuum. If people exiting an airplane landing in a warm city are observed to be wearing winter coats, the chances are good that they came from someplace cold. Similarly, if certain types of bacteria are adapted to space, there is reason to suspect they came from space. Accordingly, samples containing large varieties of bacteria could be exposed to space conditions prevailing in space. Those which survive the best could then be cultured and identified for further focused study. Evolution since arrival could have erased any past encoded genetic message, but remnants might still be found. There is only one way to find out.
But bacteria can evolve quickly, thereby potentially erasing essential information about the past of those who joined Earth’s Melting Pot some time ago. Therefore the most convincing place to look for microbial astronauts would be in space itself. Spacecraft carrying aerogels or other suitable capture media could be deployed with the mission of trying to gather spacebugs in flight. In principle they could be sent anywhere, with perhaps the most promising location being the vicinity of a comet as it outgases volatiles through its trip through the inner solar system. This could be a favorable location for space microbe collecting because it is possible that Oort Cloud objects might collect such interstellar voyagers over time, and then, when heated during close solar approach, release them in large numbers along with the vapors of the frozen volatiles that preserved and held them until that time. (Hoyle 1981). However the problem with this approach is that the characteristic relative velocities of objects moving in various orbits in space exceed several kilometers per second, so that microbes on one such trajectory slamming into anything remotely as dense as an aerogel on another would almost certainly be destroyed on impact. What is needed is an extremely diffuse medium of large expanse that can be used to slow the fast-moving microbes down to a near halt to so that they can be gathered without harm on a solid collecting surface. Fortunately, there are such media available. They are called planetary atmospheres.
Atmospheric entry from space is ordinarily thought of at a very high temperature affair, and it certainly can be. But the temperature reached will be a function of the object’s mass to surface area ratio, or ballistic coefficient. Objects like the Space Shuttle or Dragon capsule typically hit the atmosphere with ballistic coefficients between 100 and 1000 kg/m3, and these glow incandescent during reentry. However, as design studies for the German Mars Society’s ARCHIMEDES mission showed in considerable detail, a balloon with a ballistic coefficient on the order of 1 kg/m2 might well survive atmospheric entry at Mars without ever exceeding the temperature limits of Mylar. A spherical bacteria with a radius of 1 micron would have a ballistic coefficient three orders of magnitude lower still, and readily be able to avoid incineration after entry at speeds on the order of the Earth’s 11 km/s escape velocity. After being slowed down in such a manner, such a microbe would continue to fall downward, with a terminal velocity of about 3 m/s at 40 km altitude, decreasing to 1.5 m/s at 30 km altitude, and still slower at yet lower altitudes. Larger bacteria would have faster terminal velocities, with falling speed increasing as the square root of the microbe radius. For similar reasons, any microsailcraft with a low enough ballistic coefficient to escape its solar system using light pressure would have no problem safely entering the atmosphere of practically any planet.
It is possible to fly balloons at 40 km altitude, and use them to try to collect microbes. In the 1960s this was actually done by (later) Viking chief scientist Jerry Soffen, who did indeed discover viable microbes at that altitude, in such numbers that the results were considered so counterintuitive that the experiments were discontinued. (Hoyle 1981). While it is clear that any microbes found in the Earth’s upper stratosphere could be the result contamination from the biosphere below, such upward delivery by upwelling is made difficult by the strong temperature inversion in the stratosphere. This inversion places very cold (-50 C at 25 km) air below warm air (30 C at 40 km), and thereby suppresses upward convection. As a result, there is reason to suspect that microbes collected at 40 km altitude might be from space.
But how could we know? One way is by looking at the nature of the organisms collected themselves. The earliest microbes of Earth were anaerobic archaea. These can no longer live on the surface of the Earth, because they cannot tolerate the presence of oxygen. However, if life on Earth were begun via panspermia, such organisms would have been right stuff to get the ball rolling, because the surface of the prebiotic Earth was a suitable habitat for them. Furthermore, if anyone – be it ET or Nature – is still trying to seed life on prebiotic worlds today, it is these organisms, rather than the Earth’s surface biosphere currently dominant types, who would be the required pioneers. So, in short, what we should look for with our balloon-borne microbe collecting systems are anaerobes, who may exist within the Earth, but not on it, and who have no business flying around the stratosphere unless they just got off the boat.
It is also possible to fly balloons in the atmospheres of Venus and Mars. Both of these planets have sterile surfaces, effectively ruling out contamination via upwelling from below. Venus is especially attractive in this regard, as its thick atmosphere readily facilitates ballooning (two Soviet Vega balloons were successfully flown in the Venusian atmosphere in 1985) and its hot surface precludes indigenous life entirely. Any microbes collected by balloon-borne platforms floating in the atmosphere of Venus – and arguably Mars – clearly would have to come there from space, although the ultimate source of many of them might still well be the Earth.
Perhaps a particularly interesting time to conduct such experiments, whether in Earth’s atmosphere or elsewhere, might be during cometary events. If collecting balloons were flown at various times, including both normal conditions and during or shortly after cometary encounters, the difference in results could be instructive.
Using such techniques, suspects for either natural panspermia or ETI influenced microbial data transmission could be identified. The genome of these top suspects should be sequenced first, identifying which genes play a role in the functioning of the microorganism itself, and which appear to be simply along for the ride. The first set pertains to the messenger, but the second could contain the message. Perhaps it can be decoded, or if not decoded, at least examined for a format suggestive of an artificial design.
Of course, if we captured not merely microbes, but actual microsailcraft, the case for ETI initiated microbial data transmission would be settled out of hand. In that case, the key question would move directly to decryption.
If the transmitter is biological in nature, it would appear to be most logical that the receiver should be as well. If messages are being sent using genes, perhaps their meaning can best be found by inserting them in samples of terrestrial life to see what comes forth. The best terrestrial organisms to use as receivers might be bacteria themselves, as of all life here, they are most adept at adopting and putting to use new genetic information. These could be used to both receive and amplify the signal and then deliver it to more complex organisms. Perhaps novel traits could be made to appear. While radar-wielding birds are not to be expected, there are large numbers of potential animal body plans that are not currently in use of Earth. Many such plans, representing whole phyla of animal life, were briefly exhibited on our planet during the period of first flourishing of multicellular life known as the Cambrian Explosion, some 550 million years ago, only to go extinct shortly thereafter. If genes carried by suspected astrobacteria were found to induce the appearance of traits representative of such extinct phyla or other unknown animal or plant types in current life, that would be very exciting.
Of course, we might not be so lucky. A mother seeking to promote the intellectual development of her child might leave works of literature for a 16 year old, chapter books for an 8 year old, picture books for a 4 year old, and letter blocks for a 2 year old about the house in places where her child might find them. By the same logic, if ET wanted to promote evolution, he might send types of microbes adapted to successive stages of biospheric evolution containing only the information needed for the next steps, rather than the whole library of potential plans right from the start. After all, it would be futile to leave a copy of War and Peace in the nursery of a 2 year old. But even letter blocks are a dead giveaway for developmental intent. So perhaps rather than finding the genes for creating fish, trilobites, edicaria, or even eukaria in anaerobes arriving from space, we might hope to find plans for just the next step, for example chlorophyll. Any microbes carrying plans for further steps might be designed to make their way after arrival in more developed (i.e. oxygenated) phases of the biosphere, and thus be harder to identify as extraterrestrial messengers.
Still, any genetic information of a forward-looking nature carried by microbes strongly suspected of recent arrival should be a Wow signal for the ETI search. If such organisms were found to induce the appearance of such traits either uniquely or with markedly greater effectiveness than more mundane microbes, the case for evolutionary influence from space would be proven.
But there might be an even simpler way to search, because the biosphere has been acting as a giant receiver and amplifier for such messages for the past 3 billion years. That is, the history of such messages may be recorded in the biosphere itself. Consider this: the mustangs of the American west are well-adapted to their current environment, and might appear to a naïve biologist to be a product of local Darwinian evolution. But, while plausible, this conclusion would not be entirely correct. In fact, mustangs are descended from horses that escaped the Spanish Conquistadors, and their ancestors were the products of selective breeding to enable them to carry medieval knights. No doubt the genes for such past incarnations as a breed of heavily-muscled carriers of armored knights are still to be found in the cells of mustangs today, and could be activated to reveal themselves as actual traits in a mustang colt by an appropriate program. This would prove that the mustangs had a previous form that was actually a product of “intelligent design.” Furthermore, it is probably the case that if someone wanted to breed mustangs back into knight-carriers, they could do it much faster than by using horses with no such genetic history in their ancestral line. In short, both their history and some potential forward-looking traits are encoded in their genome.
This brings us back to the question of the mice and the fish. We know there are fish traits encoded in mice. No surprise there; mice evolved from fish, so naturally they carry the record of their previous evolutionary career. But do fish carry in their genome plans for any of the noteworthy traits of mice? They very well might, but perhaps only because bacteria can move genetic material around from one species to another. But if this were all there were to it, they might very well carry similar amounts of genetic material transmitted to them from species, such as insects, that are not fish descendants. However, if fish were found to be carrying genes for not only mammalian traits, but the whole roadmap of amphibians, reptiles, and mammal-like reptiles leading from fish to mice, – or even just the first essential steps on that path – that would show that somebody had been doing some serious advance planning, at least taking the trouble to shout some useful advice into the arena.
Could potential draft plans for future biospheric evolution be preprogrammed into the genes of space-traveling microbial messengers? Could the history of such past messages be recorded in the genomes of species all around us? Let’s have a look and see.
Predictions and Conclusions
To be useful, any scientific theory needs to be testable and falsifiable. While the balloon borne microbe or microsailcraft collection experiments described above could potentially offer strong evidence in favor of panspermia, they cannot disprove it because a negative result could be explained away by the argument that the flux of microbes from space is simply too low to be detected by such means. Similarly, the genomic search for forward-looking traits in terrestrial organisms could reveal directed panspermia, but not disprove it. Such experiments should be done regardless because they are cheap and might produce profound results. But if they are unsuccessful in producing useful data, a more muscular approach to settling the matter will be necessary. This can be done through the exploration of Mars.
Mars was once a warm and wet planet, which could have hosted life on its surface, and there is strong evidence that there is still liquid water to be found underground on Mars, which could serve as a habitable environment for microbial life today. If there ever was life on the surface of Mars, it is reasonable to assume that it retreated into the groundwater when conditions on the surface of the planet deteriorated, much as the anaerobic archaea did on Earth after the oxygenation of the atmosphere made the surface here inhospitable for their kind. If we could go to Mars and sample the groundwater, what we find, or fail to find, therein would be very informative. There are four primary possibilities.
1. We find no life in Mars groundwater. This is a very unlikely result, because Mars almost certainly had life on its surface at one time, if from no other source than the Earth. But if that should be the finding, it would prove the case for not only geospermia, but unique geospermia, occurring on Earth but not on the similar early Mars, indicating that life is rare in the universe.
2. We find life in Mars groundwater, which uses the same biochemistry as Earth life, but including more primitive free living representatives ancestral to bacteria. This would refute panspermia as an origin of life theory, instead showing that life on Earth originated on Mars. It would also support the conjecture that life is common in the universe, as apparently it could evolve from chemistry readily on a primitive terrestrial type planet.
3. We find life in Mars groundwater which uses a different biochemistry than what we find on Earth, i.e. a second genesis. This would refute natural panspermia, but prove that life is very common and quite diverse in the universe, as it would be seen that life could originate from chemistry, de novo, two different ways, on two out of two typical primitive terrestrial planets.
4. We could find life in the groundwater of Mars that uses the same biochemistry as Earth life, manifesting similar bacteria forms, with no more primitive free-living representatives evident. This is what natural panspermia would predict. It may be argued that the same result could be achieved by the origin of life on either Earth or Mars, with subsequent transfer between them as well as extinction of the ancestral forms on both worlds. But this means that the current alibi of the geospermians – “the origin did happen here, really it did, we believe that sincerely, even though experiments show that conditions here were unfavorable, we’ve just lost all the evidence” – would need to be stretched to two worlds.
It should be noted that while operation of robotic rovers on the surface of Mars might serve to falsify alternative 1 (which is fantastical in any case as it requires accepting the conceit that not only is Earth uniquely capable of originating life, but that life from Earth cannot spread to other habitable places nearby) by discovering fossils, it cannot affirm it. More importantly, such robotic exploration techniques would be incapable of distinguishing between alternatives 2,3, or 4, which is the most critical scientific question. Resolving between these alternatives, which have profound implications for the nature of life and the universe, will require drilling to depths on the order of a kilometer to reach groundwater, bringing up samples, culturing them, and subjecting them to biological analysis. From a practical point of view, this can only be done by sending human explorers to Mars.
But which of these alternatives is most likely? I predict that number 4 is what we will find. The reason is simply this: The Milky Way galaxy predates the Earth by 8 billion years. The early Earth was not exceptional in any significant way, so that if life could evolve here, it could have evolved first on hundreds of billions of other possible locations. Furthermore, the first life that did so which developed adaptations allowing it to survive interstellar flight would necessarily spread throughout the galaxy in less than a billion years by natural collisional processes. This would result in the appearance of life on any planet as soon as conditions there were suitable, which is exactly what we observe in the fossil record on Earth.
For these reasons I believe that natural panspermia is extremely probable, and that the results of Mars exploration will prove to be consistent with it. But what about directed panspermia? While the history of life on Earth has demonstrated the ability of biospheres to evolve increasingly intelligent species (Morell, 2013), what reason is there to believe that such intelligent extraterrestrials should want to spread their kind around? None, except this: If there were any such species, it would be their kind that would get spread around. If we find any forward-looking traits encoded in the Martian biota, their handiwork would be there for all to see.
Finally, it may be observed that a program of directed panspermia using microsailcraft is well within humanity’s current technological means. If it turns out that no one else has been doing the noble work of spreading life throughout the universe, perhaps we should get the ball rolling ourselves.
Acknowledgement
I wish to thank Chris McKay of NASA Ames Research Center and Paul Davies of Arizona State University for useful comments on early drafts of this manuscript.
References
Arrhenius, S. (1908) Worlds in the Making: The Evolution of the Universe, Harper and Brothers Publishers, New York and London, 1908
Ayre, J. (2012)”Text of a full book encoded in DNA,” Planetsave August 18, 2012, http://planetsave.com/2012/08/18/text-of-a-full-book-encoded-in-dna/ Accessed Feb 25, 2017.
Bernhardi, F. v. (1912) Germany and the Next War, 1912, Kindle edition May 2012 http://www.goodreads.com/book/show/19347632-germany-and-the-next-war Accessed February 26, 2017
Bhuler B.-A. et al (2015) “A Molecular Mechanism for the origin of a key evolutionary innovation, the bird beak and palate, revealed by an integrative approach to major transitions in vertebrate history,” Evolution, June 30, 2015, http://onlinelibrary.wiley.com/doi/10.1111/evo.12684/abstract;jsessionid=A0CA3A47A57D5319E64A23C1ECA41B02.f02t03 Accessed February 27, 2017
Cocconi, G and P.Morrison (1959) “Searching for Interstellar Communications,” Nature, vol 184, no 4690, pp. 844-846, September 19, 1959. http://www.bigear.org/vol1no1/interste.htm Accessed February 26, 2017
Davies, P. (2010) The Eerie Silence: Renewing Our Search for Alien Intelligence, Mariner Books, 2010.
Drake, N. (2017) “This May Be the Oldest Known Sign of life on Earth,” National Geographic, March 1, 2017 http://news.nationalgeographic.com/2017/03/oldest-life-earth-iron-fossils-canada-vents-science/ Accessed March 12, 2017.
Griebel, H. et al (2004) “Project Archimedes: A novel approach to balloon deployment on Mars” 55th International Astronautical congress, Vancouver, CA, 2004 IAC-04-Q.P.02 https://arc.aiaa.org/doi/abs/10.2514/6.IAC-04-Q.P.02
Herkewitz, W. (2016) “Scientists turn bacteria into living hard drives,” Popular Science, June 9, 2016 http://www.popularmechanics.com/science/animals/a21268/scientists-turn-bacteria-into-living-hard-drives/ Accessed February 25, 2017
Hitler. A. (1941) “The law of existence requires uninterrupted killing, so that the better may live,” private communication to associates made while issuing the order to exterminate the population of Leningrad, September 1941.
Hogenboom, M. (2015) “Chicken Grows Face of Dinosaur,” BBC News, May 13, 2015 http://www.bbc.com/earth/story/20150512-bird-grows-face-of-dinosaur Accessed February 27, 2017
Hornek, G., D. Klaus, and R. Mancinelli, (2010) “Space Mircobiology,” Microbiology and Molecular Biology Reviews, 2010 Mar, 74(1): 121-156.
Hotopp, J.D. (2011) “Horizontal Gene Transfer Between Bacteria and Animals,” Trends Genet. Feb. 18, 2011 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3068243/ Accessed February 27, 2017
Hoyle, F. and C. Wickramasinghe, (1981) Evolution from Space, Simon and Schuster, New York, 1981
Margulis L. and D. Sagan, (2008) Acquiring Genomes: A Theory of the Origin of Species, Basic Books, 2008
Melosh, J (1988). “The rocky road to panspermia.” Nature 332:687-688.
McDowell, N. (2003) “Data Stored in multiplying Bacteria,” New Scientist, January 8, 2003, https://www.newscientist.com/article/dn3243-data-stored-in-multiplying-bacteria/ Accessed February 25, 2017.
Morell, V. (2013) Animal Wise: How We Know Animals Think and Feel, Crown, New York, 2013. A very instructive refutation of the thesis that intelligence is qualitative phenomenon singular to humans, as opposed to a quantitative phenomenon that has been developing in terrestrial animals over time.
Service, R. (2017) “DNA Could Store All the World’s Data in One Room'” Science, March 2, 2017,
http://www.sciencemag.org/news/2017/03/dna-could-store-all-worlds-data-one-room. Accessed March 12, 2017.
Wilkins, A. (2010) “Bioencryption can store almost a million gigabytes of data inside of bacteria,” IO9, November 26, 2010 http://io9.gizmodo.com/5699767/bioencryption-can-store-almost-a-million-gigabytes-of-data-inside-bacteria Accessed February 25, 2017.
Yong, E. (2016) “I contain Multitudes: The Microbes within Us and a Grander view of Life,” Ecco, 2016
Zubrin, R. (2001) “Interstellar Panspermia Reconsidered,” Journal of the British Interplanetary Society Vol 54, pp. 262-269, 2001, http://www.jbis.org.uk/paper.php?p=2001.54.262, Accessed February 26, 2017
Zubrin, R and D. Andrews, (1989) “Magnetic Sails and Interplanetary Travel,” AIAA 89-2441, AIAA-ASME Joint Propulsion Conference, Monterey, CA, July 10-12, 1989. Reprinted in Journal of Rockets and Spaceflight, April, 1991.



How to keep humans from ruining the search for life on Mars
Microbes from Earth might complicate human junkets to the Red Planet
By Lisa Grossman
11:30am, January 10, 2018
https://www.sciencenews.org/article/manned-mission-mars-humans-microbes-life
Martian Methane Varies with the Seasons
And microbial activity could be the reason.
By Dirk Schulze-Makuch
airspacemag.com
3 hours ago
https://www.airspacemag.com/daily-planet/martian-methane-varies-seasons-180967892/
To quote:
Several explanations have been advanced for these observations. Some scientists think the methane comes from biological sources, either from still living microbes, or as a remnant signature of life processes from the planet’s wetter ancient past. If they’re correct, that would mean life has existed on Mars, and probably still does today, in some environmental niches. But how to prove this, short of a dedicated life-detection mission? A variety of inorganic sources have also been proposed for the methane, but the challenge there is to reach the levels of concentration found in the Martian atmosphere, which translates to thousands of tons.
The new observation of seasonality supports the biological hypotheses. Life on Earth has a distinct seasonal signature, thriving in the warm summer months and going dormant in the winter months, particularly in the colder latitudes. The same should be true for Mars. There are also chemical processes that exhibit seasonal variations, but according to Webster and his colleagues, the observed signal is about three times greater than would be the combined effect of these inorganic processes (such as methane production from ultraviolet degradation of meteoritic material, or sublimation and freezing of carbon dioxide at the poles).
The previous discovery of methane plumes already hinted toward biology, and the seasonality of the methane concentrations observed over the last five years (2.5 Martian years) only strengthens that possibility. However, more evidence is needed. And that we might get with ESA’s ExoMars Trace Gas Orbiter which is expected to begin its study of the Martian atmosphere this spring.
Is the origin of life just cosmic dust in the wind?
January 23, 2018
by Karl Gruber
“The cosmos is within us. We’re made of star stuff.” Thanks to a new study, this famous phrase by iconic astronomer Carl Sagan, now has some more support.
Just like the wind blows dust into your house, cosmic dust can be blown across space and between planets. Cosmic dust is the stuff planets and stars are made of: carbon, oxygen, iron and other tiny particles less than 1 micrometre in size.
A new study proposes that life on Earth started from particles flying as dust from outer space.
Full article here:
https://phys.org/news/2018-01-life-cosmic.html
Overall, a very well thought out article. I enjoyed the implications drawn from the SETI data we have available, and your analysis of both the potential consequences if the mechanism for interstellar communication is true, but also how/if it is not.
I believe simple ‘interstellar-ready’ shaping of the bacteria can greatly ease travel for thousands of years.
Faster initial velocity can come from an electrostatic sail configuration, with a large flat negatively-charged disk allowing >500km/s boost from solar wind.
DNA density alone is 2000kg/m^3. The bacteria will likely be dry spores, so total density closer to 2x the value you used (1000kg/m^3).
As mentioned above, disk shapes can greatly improve initial velocity, then turns into cylinder to reduce cross-section during transit.
An opaque shell can vastly reduce exposure to UV during transit. Cylinder with DNA in the back can provide some protection from collisions over time.
Bacteria can be used to broadcast information… but also provide instructions on how to set up the much more energy intensive two-way communications through radio.
http://astrobiology.com/2018/01/implications-of-captured-interstellar-objects-for-panspermia-and-extraterrestrial-life.html
Implications of Captured Interstellar Objects for Panspermia and Extraterrestrial Life
Press Release – Source: astro-ph.EP
Posted January 31, 2018 8:32 PM
We estimate the capture rate of interstellar objects by means of three-body gravitational interactions. We apply this model to the Sun-Jupiter system and the Alpha Centauri A\B binary system, and find that the radius of the largest captured object is a few tens of km and Earth-sized respectively.
We explore the implications of our model for the transfer of life by means of rocky material. The interstellar comets captured by the “fishing net” of the Solar system can be potentially distinguished by their differing ratios of oxygen isotopes through high-resolution spectroscopy of water vapor in their tails.
Manasvi Lingam, Abraham Loeb
(Submitted on 30 Jan 2018)
Comments: 9 pages; 0 figures
Subjects: Earth and Planetary Astrophysics (astro-ph.EP)
Cite as: arXiv:1801.10254 [astro-ph.EP] (or arXiv:1801.10254v1 [astro-ph.EP] for this version)
Submission history
From: Manasvi Lingam
[v1] Tue, 30 Jan 2018 23:46:23 GMT (71kb)
https://arxiv.org/abs/1801.10254
Astrobiology
http://astrobiology.com/2018/02/dynamical-and-biological-panspermia-constraints-within-multi-planet-exosystems.html
Dynamical and Biological Panspermia Constraints Within Multi-planet Exosystems
Press Release – Source: astro-ph.EP
Posted February 13, 2018 9:57 PM
As discoveries of multiple planets in the habitable zone of their parent star mount, developing analytical techniques to quantify extrasolar intra-system panspermia will become increasingly important.
Here, we provide user-friendly prescriptions that describe the asteroid impact characteristics which would be necessary to transport life both inwards and outwards within these systems within a single framework. Our focus is on projectile generation and delivery and our expressions are algebraic, eliminating the need for the solution of differential equations. We derive a probability distribution function for life-bearing debris to reach a planetary orbit, and describe the survival of micro-organisms during planetary ejection, their journey through interplanetary space, and atmospheric entry.
Dimitri Veras, David J. Armstrong, James A. Blake, Jose F. Gutierrez-Marcos, Alan P. Jackson, Hendrik Schaefer
(Submitted on 12 Feb 2018)
Comments: Accepted for publication in Astrobiology
Subjects: Earth and Planetary Astrophysics (astro-ph.EP); Solar and Stellar Astrophysics (astro-ph.SR)
Cite as: arXiv:1802.04279 [astro-ph.EP] (or arXiv:1802.04279v1 [astro-ph.EP] for this version)
Submission history
From: Dimitri Veras
[v1] Mon, 12 Feb 2018 19:00:03 GMT (1850kb)
https://arxiv.org/abs/1802.04279
Astrobiology
Your comment on possible hidden messages in junk DNA. Please be aware that DNA is composed by nucleic acids and not by amino acids. There is some difference between proteins and DNA.
We would have been forbidden to even undergo a next examination after such a blunder.
Cause of Cambrian Explosion – Terrestrial or Cosmic?
https://www.sciencedirect.com/science/article/pii/S0079610718300798
Our DNA Might Be the Most Valuable Thing in a Trade Deal With Aliens
Or, how to avoid a “Mars Attacks”-style bloodbath when we eventually encounter extraterrestrials.
Mirjam Guesgen
Jun 13 2018, 10:41am
https://motherboard.vice.com/en_us/article/kzkk5w/earth-offer-dna-in-alien-trade-deal-meti-seti
To quote:
Trading DNA comes with the added benefit of preserving genetic information elsewhere, said Helman. It’s like a conservation insurance policy for Earth, in case we wipe out life all by ourselves through nuclear war or climate change. The possibility of a civilizational insurance policy might also drive people to try harder to conserve creatures now, reasoned Helman.
“You can’t form the basis for trade if you don’t have the goods,” he told me. “If we’re destroying biodiversity then we’re limiting the unique goods we have. It’s a utilitarian reason to promote biodiversity.”
A Simple Chemical Process May Have Led To The Origin Of Life On Earth
Press Release – Source: Tokyo Institute of Technology
Posted June 13, 2018 7:44 PM
Research has shown that reactions of alpha-hydroxy acids, similar to the alpha-amino acids that make up modern proteins, form large polymers easily under conditions presumed prevalent on early Earth.
These alpha-hydroxy acid polymers may have aided in the formation of living systems on early Earth.
There are different theories for how life first formed on early Earth. One popular one suggests that life may have arisen in specialized environments, such as tidal pools or shallow water hot springs, where simple chemical reactions would have helped generate life’s precursors. All life is made up of polymers, large molecules made up of a sequence of molecules called monomers. A key question is how biological polymers could have formed without enzymes on early Earth.
While environments of early Earth might have had monomers that could give rise to life, it would have been difficult for polymers to arise from these without the help of enzymes. In this case, the team showed these polymers could have formed with alpha-hydroxy acids before the existence of enzymes on early Earth.
This multi-national team showed that hydroxy acids polymerize more easily than amino acids, and that they could have provided the necessary toolkit to kick-start the formation of more complex molecules for the origin of life on Earth.
To simulate various primitive environments, the team reacted alpha hydroxy acids under varied conditions of pH and temperature, from room temperature to boiling hot. Using sophisticated high-resolution mass spectrometry and data analysis software, they showed that these polymers can form over a range of conditions.
The team showed further, that using a mixture of different types of alpha-hydroxy acid that they could form huge numbers of types of polymers, in effect creating vast ‘libraries’ of different chemical structures, which can remain stable over a range of environmental conditions. The work culminated with the creation of a polyester library made up of five kinds of alpha hydroxy acids (Figure 1). They showed that this simple-to-create library could contain hundreds of trillions of distinct polymer sequences.
This research opens new areas of exploration of the chemistry that may have led to the formation of life. The authors are keen to extend this work by investigating the types of functions of alpha-hydroxy acid polymers might be able to carry out, and other types of simple chemicals that could interact with these, possibly leading to self-replicating chemical systems.
http://astrobiology.com/2018/06/a-simple-chemical-process-may-have-led-to-the-origin-of-life-on-earth.html