In our continuing look at biosignatures that could flag the presence of life on other worlds, we’ve sometimes considered the so-called ‘red edge,’ the sharp change in reflectance of vegetation that shows up in the near-infrared. It’s worth remembering that vegetation is the largest reflecting surface on Earth (about 60 percent of the land surface), with an increase in reflectance that shows up around 700 nm. As Alex Tolley explains below, the red edge may shift depending on the evolution of plant life and the variables, including light intensity but comprising many other factors, that would affect life on M-dwarf planets. These are the first whose atmospheres we’ll be seriously examining for biosignatures, and the question of how to extrapolate from Earth life to environments as exotic as these is complex. A Centauri Dreams regular, Alex reminds us that vegetative life may prove adaptable in ways that will surprise us.
by Alex Tolley

Artist’s conception of Proxima Centauri b. Credit: M. Kommesser
Xi Jinping University, New Beijing, Mars
NEWS RELEASE
FOR IMMEDIATE RELEASE
Contact Wendy Ho, Media Liaison.
wendy.ho@univxijinping.co.mars
Nov. 11, 2091
The first image of the surface ofProxima b is released today. Captured by the solar focal telescope (SoFoT), the 2-megapixel image captures the 3?4 illuminated planet’s surface. This is the first direct image of the planet that does not rely on using the EPSI inversion model for the reflected solar radiation. Both continents and oceans are clearly visible. The ocean appears wine-dark, and the continental landmasses are partially covered in a purplish color that is most probably vegetation. This vegetation contrasts with the orange-colored material of the deserts. There are no polar ice caps. The vegetated areas reach into the very high latitudes of both poles. The previous spectroscopic analysis had indicated that the atmosphere contained the biosignature gases oxygen and methane. The likely presence of vegetation suggests the source of this oxygen is photosynthesis. The color of the vegetation is likely due to the shift of the maximal light absorption into the red and near-infrared, as well as the lower flux of blue and green wavelengths of light in Proxima’s spectrum. This gives an orange cast to the landscape, with the bluer color of the local chlorophyll analog dimmed. The color difference seen is similar to that of undersea life on Earth which looks dark in the blue light at depth, but which are brightly colored in white light.
Professor Zhang Yong suggested that the photic zone in the oceans was likely quite shallow due to the low blue light emission of the star and that possibly the rate of photosynthesis was lower than on Earth. The Chinese star probe, New Dawn, is currently on its way to Proxima. Launched in 2077 it is expected to reach the Proxima system in 2099. The image of Proxima b is the first to have life confirmed visually and indicates that the probe will confirm life on the planet. The IAU will meet in December to select an official name for the planet and its features.
The abundance of M-dwarfs in the galaxy, as well as the M-dwarf, Proxima Centauri, having a rocky planet in the habitable zone (HZ) has renewed speculation of what adaptations photosynthetic plants might have to the red-shifted spectrum of these stars. Figure 1 below shows the difference in spectrum received by Earth compared to that of Proxima b. The red-shifted peak emission of Proxima is evident, as well as the high intensity of extremely short wavelength radiation compared to Earth.
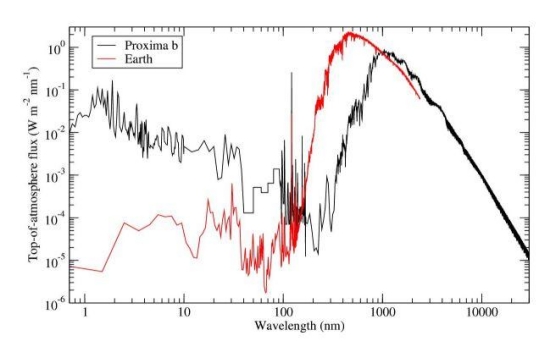
Figure 1. Top-of-atmosphere full spectral irradiance received by Proxima b (black) and the Earth (red). An orbital distance of 0.0485 AU is assumed for Proxima b. [4]
Most speculation about the adaptation of plants concerns multicellular, terrestrial green plants that have clothed the surface of our continents. These plants are almost all green plants that contain chlorophylls a and b, whose peak absorption wavelengths are both in the blue and red end of the spectrum, and therefore reflecting wavelengths between these peaks, that we perceive as green (see figure 2). The adaptation of green plants to our sun’s spectrum leads to the speculation that plants on exoplanets around M-dwarfs will similarly adapt, with a dominant absorption that would extract more energy from the red end of the spectrum and therefore changing the apparent color of their leaves. Some have speculated that the absorption of more of the light spectrum that we are adapted to see would render the leaves almost black to our eyes.
Jack O’Malley-James, School of Physics and Astronomy, University of St Andrews:
“Plants with dim red dwarf suns for example, may appear black to our eyes, absorbing across the entire visible wavelength range in order to use as much of the available light as possible.”
In this essay, I will argue that there is evidence that chlorophylls can evolve to trap much redder light, but that this is too simplistic a story when considering how plants may evolve in response to the spectrum of M-dwarfs like Proxima.
While green plants dominate the Earth today, there are other plants that use different combinations of chlorophylls. The red algae (Rhodophyta) use chlorophylls a and d, plus a red-colored accessory pigment, phycoerythrin. These ensure that the dominant absorption is at the blue end of the spectrum, and is adapted for their deeper marine habitats where the penetrating sunlight is blue filtered. Without white light to illuminate these algae, they appear black to our eyes. The brown algae, which includes the kelps, use chlorophylls a and c and an Accessory pigment, fucoxanthin. Again, the peak absorption wavelengths are at the blue end of the spectrum as befits their subsurface, marine habitats. All these plants, whether multicellular or unicellular, capture the energy of the sun to fix carbon via photosynthesis. This reduces the carbon of the carbon dioxide (CO2) with the hydrogen in water (H2O) to produce sugars and release oxygen (O2) as a byproduct. However, this is not the only method of photosynthesis.
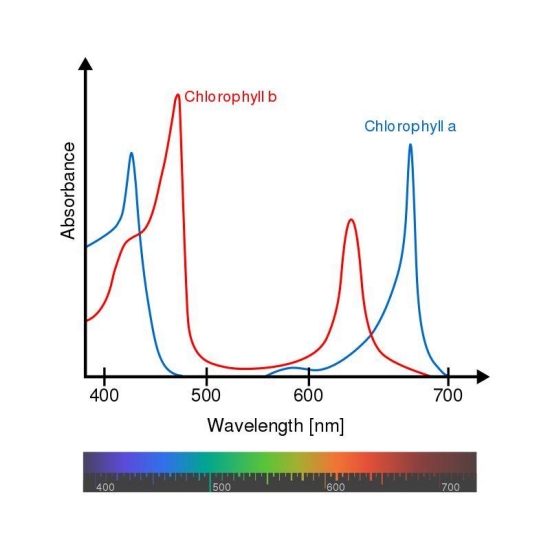
Figure 2. The absorption spectra of chlorophylls a and b, found in green plants and green Algae.
Bacteria also trap electromagnetic radiation and fix carbon. However, they use either hydrogen gas (H2) or hydrogen sulfide (H2S). In the latter case, they release particulate sulfur. Both of these types of photosynthesis require anoxic conditions and the bacteria live in hot springs or vents where these 2 gases are available. Both plant and bacterial chlorophylls are based on tetrapyrrole molecules. The absorption peaks of which are changed by small modifications of the structure.
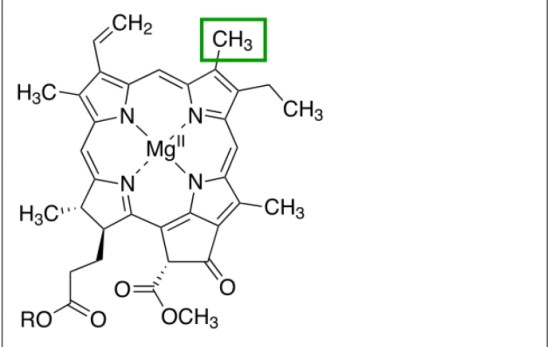
Figure 3. Tetrapyrrole ring for chlorophyll a.
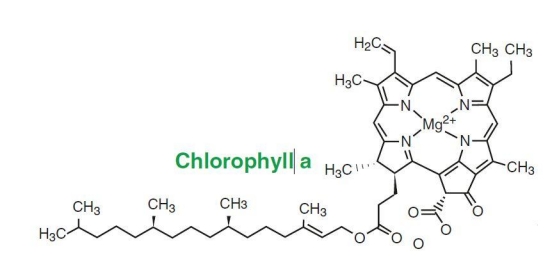
Figure 4. The structure of chlorophyll a. This is the primary photosynthetic chlorophyll.
While terrestrial life has evolved under our sun, there is the question of whether these light-absorbing molecules can evolve to meet the different spectra of other stars. While there is interest in whether planets in the HZ of M-dwarfs can even evolve life, that life would be limited to low energy, unicellular chemotrophs unless they can harvest the longer wavelength energy of their star.
The first question is whether these core, tetrapyrrole, light-absorbing molecules have already evolved on Earth to trap longer wavelength light. The obvious place to look is with the bacteria that live in very low light environments yet still photosynthesize to fix carbon.
The answer is yes. Of the 8 known bacteriochlorophylls, bacteriochlorophyll b that is found in purple bacteria has peak absorption wavelengths in the infrared (IR) of 835–850nm and 1020–1040nm.
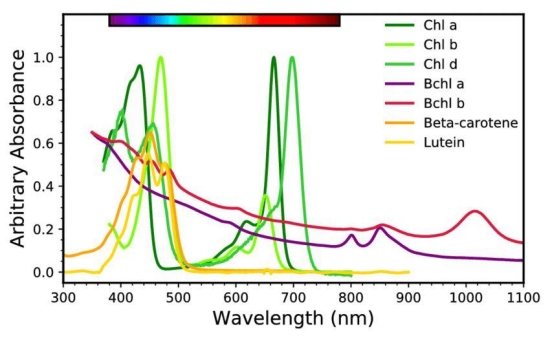
Figure 5. Photosynthetic and other light-absorbing pigments. Absorbance spectra of selected chlorophylls (Chls), bacteriochlorophylls (Bchls), and carotenoids showing the wavelengths of absorption peaks. A visible spectrum colorbar is shown at the top. The y-axis scaling is arbitrary. The Chl absorbance data are of extracted pigments in methanol solution from Chen & Blankenship (2011). The Bchl a and Bchl b data are of whole cells and originate from Cogdell & van Grondelle (2003). The beta-carotene (carotenoid) spectrum is of pigment extract in hexane from Dixon et al. (2005), while the lutein (carotenoid) is from Janik et al. (2008). Pigments dissolved in solvents have absorption peaks slightly shifted from those in cells. These data are publicly available on the Virtual Planetary Laboratory Biological Pigments Database (http://vplapps.astro.washington.edu/pigments). [3]
Relatively recent work by Faries [1] and ? Vairaprakash [2] on bacteriochlorins, modifying their side groups has shown that the peak absorption wavelength can be pushed out into the near infra-red. The number of variants in the chlorin ring exceeds that found in nature, and shows how small changes can modify the peak absorption wavelength. Figure 6 shows the results of these experiments that aimed to understand exactly how to tailor these molecules for different peak absorptions, primarily for industrial purposes. Note that all the absorption spectra retain their double peaks at each end of the visible spectrum. It should also be noted that there is clearly a drift towards a secondary peak at the short wavelengths that get shorter as the maximum peak wavelength gets longer, however this appears bound by a lower limit around 360nm, near the violet to ultraviolet transition. Whether these modifications could be produced by natural biochemistry would be interesting. The reactions needed may be outside of the possible repertoire, or too complex to prove evolutionarily viable.
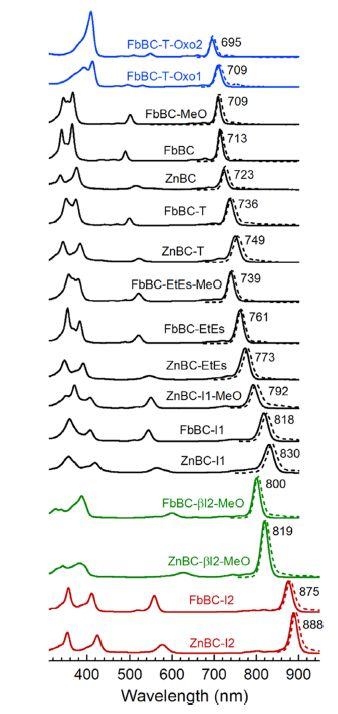
Figure 6. The expansion of the spectral window for the bacteriochlorins by placement of uxochromes at designated sites affords a commensurate increase in the photochemical tailorability of this class of nature-inspired molecules. Collectively, the results provide fundamental insights into the rational design and synthesis of red- and NIR-absorbing bacteriochlorins for solar-energy and life sciences research. [2]
This bodes well for the possible evolution of photosynthesis on planets around M-dwarfs. It indicates that life could evolve to harvest energy not just of M-dwarfs, but even brown dwarfs. This would at least ensure that non-oxygenic photosynthesis by bacteria is possible to fix Carbon.
Photosynthesis in green plants consists of 2 systems, photosystem I (PS I) and photosystem 2 (PS II). PS I is the earlier system to evolve and is the reaction that drives carbon fixation in nonoxygenic, photosynthetic bacteria. PS II evolved later and uses the addition of shorter wavelength, higher energy light, captured more by chlorophyll b to split the H2O molecule to liberate O2 as a byproduct before the reactions to fix carbon.
Experimental work [5] shows that some blue light is needed for photosynthesis to operate efficiently, although red light alone will allow suboptimal photosynthesis to occur. This is important, as it implies that even terrestrial green plants should be able to grow on an exoplanet in the HZ around an M-dwarf.
So far the story suggests that photosynthesis, both oxygenic and nonoxygenic (in bacteria) should be possible under M-dwarf light. The evolution of longer wavelength absorbing light in bacterial chlorophylls, both natural and experimentally, suggests that light trapping tetrapyrrole molecules or possible analogs, should allow the evolution of photosynthesis under the light spectrum of M-dwarfs.
While O’Malley-James (see earlier quote) speculated that plants on M-dwarf planets might use more chlorophylls to capture light at different peak wavelengths, we do not see such adaptations on Earth, even though plants live in varied light condition environments. All plants seem to produce just 2 chlorophylls, with chlorophyll a being the dominant one for photosynthesis.
The question is “Why don’t plants produce more than 2 chlorophylls?” The answer lies most probably in the cost of a larger genome to code for the extra pathways and enzymes to manufacture the chlorophylls and regulate them. Instead, plants have opted for simpler, less costly solutions. The brown algae encode the biochemistry to manufacture the accessory pigment fucoxanthin, and the red algae, the protein phycoerythrin as an accessory pigment. These seem to be a tradeoff that optimizes the capture of available light energy for growth and reproduction.
However, light-capturing molecules are not the whole story. Terrestrial, multicellular plants adapt to low light conditions by a number of strategies. These include:
1. Increasing the number of chloroplasts in their cells. This is why plants in the low light understory plants of tropical jungles tend to have dark green leaves.
2. Rather than expending energy creating lignin-rich trunks to raise the leaves of trees to the sunlight, vines simply raise their leaves by climbing those trunks, or other surfaces to reach more sunlight.
One of the main objections to life on M-dwarf exoplanets is the high ultraviolet (UV) flux. On Earth, the bacterium Deinococcus radiodurans has evolved robust biology to resist radiation damage. Other organisms might respond by remaining at depth in the oceans, or under the ice to avoid the UV intensity. If the UV is inconstant, they may adapt by other means to reduce exposure. One means is to stay unicellular and attach to a motile animal than can move to safety. On Earth, we have marine slugs that ingest algae to extract their chloroplasts for photosynthesis. Maintaining algal pouches like squid do for luminescent bacteria is also a possibility.

Figure 7. The sea slug, Elysia chlorotica, eats green algae. It retains the chloroplasts so that it can harness photosynthesis for extra energy.
It is often suggested that flares will strip the atmosphere off an M-dwarf planet. If life has evolved, atmosphere stripping would freeze the oceans, creating a thick ice crust. Photosynthetic life might be able to live below that crust, trapping the light that penetrated the icy crust, or even within the ice using the tiny liquid water fraction to allow metabolism. The conclusion I draw from our one example of Earth’s plant life, is that we should be careful in extending simple logic to speculate how plants might evolve on M-dwarf planets. Lifeforms optimize a myriad of variables to gain an edge, using different strategies to maximize gene replication. As plant breeders have long since learned, it is often not possible to optimize for one feature, such as increased seed size and production, without needing to alter other features. Light intensity and spectrum are not the only variables plants will experience on M-dwarf planets and plants will find a number of ways to adapt to those conditions, probably in ways we cannot foresee based on our experience.
A final note on detection. Because of the dominance of chlorophyll a on Earth, the reflectance of the Earth has a sharp increase at wavelengths just beyond its peak absorption. This is known as the “red edge” and is suggested as a possible biosignature. O’Malley-James and Kaltenegger [6] suggest that since the early photosynthetic cyanobacteria also have the same chlorophyll type, that the red edge signature could be used even for exoplanets at a much earlier stage of the evolution of life. However, the longer wavelength peak absorption of bacteria using different bacteriochlorophylls, and the possible evolution of longer wavelength absorbing chlorophylls suggests that this red edge may shift, perhaps into the IR. If so, some flexibility in the determination of any red edge biosignature should be entertained.
References
1. Faries, K. M., Diers, J. R., Springer, J. W., Yang, E., Ptaszek, M., Lahaye, D., … Holten, D. (2015). “Photophysical Properties and Electronic Structure of Chlorin-Imides: Bridging the Gap between Chlorins and Bacteriochlorins.” The Journal of Physical Chemistry B, 119 (24), 7503-7515. doi:10.1021/jp511257w.
2. Vairaprakash, P., Yang, E., Sahin, T., Taniguchi, M., Krayer, M., Diers, J. R., … Holten, D. (2015). “Extending the Short and Long Wavelength Limits of Bacteriochlorin Near-Infrared Absorption via Dioxo- and Bisimide-Functionalization,”? The Journal of Physical Chemistry B Vol. 119 (12), 4382-4395. doi:10.1021/jp512818g
3. Schwieterman, Edward. (2018). “Surface and Temporal Biosignatures.”
10.1007/978-3-319-30648-3_69-1.
4. Ribas et al., “The full spectral radiative properties of Proxima Centauri,” Astronomy & Astrophysics Vol. 603, A58 (July, 2017).
https://www.aanda.org/articles/aa/full_html/2017/07/aa30582-17/aa30582-17.html
5. Hogewoning, S. W., Trouwborst, G., Maljaars, H., Poorter, H., Van Ieperen, W., & Harbinson, J. (2010). “Blue light dose-responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light,” Journal of Experimental Botany, 61 (11), 3107-3117. doi:10.1093/jxb/erq132
6. O’Malley-James, J. T., & Kaltenegger, L. (2019). “Expanding the Timeline for Earth’s Photosynthetic Red Edge Biosignature,”. The Astrophysical Journal, Vol. 879 (2), L20. doi:10.3847/2041-8213/ab2769
7. Berdyugina, S. V., & Kuhn, J. R. (2019). “Surface Imaging of Proxima b and Other Exoplanets: Albedo Maps, Biosignatures, and Technosignatures,” The Astronomical Journal Vol. 158 (6), 246. doi:10.3847/1538-3881/ab2df3
8. Stevenson, D. S. (2017). Life Jim, But Not as We Know It. In The Nature of Life and Its Potential to Survive (Springer), 193–236. doi:10.1007/978-3-319-52911-0_5



Thanks for the great snowy-Monday-morning read! While Alex’s article gives an excellent explanation of what potential biosignatures from Proxima Centauri b could look like, there are hazards relying on such observations as proof of alien life on an a distant world using new observation techniques:
https://www.drewexmachina.com/2014/10/05/a-cautionary-tale-of-extraterrestrial-chlorophyll/
That article was a good read. I am also reminded how von Braun got it wrong about the density of the Martian Atmosphere. His book, “Project Mars: A Technical Tale” has a chapter (ch 4) where the density of the atmosphere of Mars is discussed. Von Braun calculated that the pressure is perhaps 1/12th that of Earth The logic based on known observations is good, but unfortunately, the observations were wrong. Because of this, if a real project had gone according to the book’s plan, it would have been a disaster as the proposed landing ships had wings to glide in the Martian atmosphere. At least SpaceX has the real data for testing their Starship as a Mars colony ship that will use aerobraking and a glide descent.
Perhaps I am thinking too much of the more modern Space Age era, but wouldn’t they have sent at least a few automated probes to Mars first to check things out?
Sending an entire manned mission all the way to the Red Planet without at least one advance scouting mission seems suicidal, I don’t care how tough and manly they were in the 1950s.
Here is von Braun’s 1950s Mars plan online:
https://www.wlym.com/archive/oakland/docs/MarsProject.pdf
Here is his plan from 1969 for landing humans on Mars in 1981. Had me and many others thinking it was actually going to happen then…
https://www.nasa.gov/sites/default/files/atoms/files/19690804_manned_mars_landing_presentation_to_the_space_task_group_by_dr._wernher_von_braun.pdf
Thank you alex for mentioning EPSI. I hope you are off by 11 years or so, and that the purple hills of Proxima b come into sharp focus in 2080 when hundreds upon hundreds of Breakthrough Starshot mini lightsails zip past at 20% lightspeed and image the entire illuminated surface. This is also why EPSI is so important. If you remember, it took weeks for the Huygens imaging team to piece together a composit picture of the landing site and its immediate environs. Imagine the challenge the Breakthrough Starshot imaging team will have potting together a similar construct of Proxima b as a result of a complete inability to point precisely at surface targets due to the OVERWHELMING SPEED at which the lightsails will be traveling. If EPSI can provide even a realistic TEMPLATE of the surface, it will make this extremely daunting task a whole lot easier. NOW: If ALL OF THE TECHNOLOGICAL ISSUES facing the Terrascope are resolved at incredible speed, you and I have an outside shot of seeing these purple hills in our lifetime. KEEP YOUR FINGERS CROSSED!
It was your comment about ESPI in a previous post that required a last draft revision. ;) I was aware of earlier work, but that paper seems to have moved the technology forward. Assuming it can be applied to exoplanets, especially rocky worlds with distinct continents and oceans, that would be very useful information to distinguish such worlds from dessicated or water worlds.
Centauri Dreams readers: To view the comment mentioned by Alex, above and all of the replies it generated, go back up to “Recent Posts”, click on “Axial Tilt, Habitability, and Centauri B.”, scroll down to “26 Comments.”, click on, and scroll down to near the bottom.
Alex, Great article, well written and researched. The view that you followed was well thought out and had me anticipating what would come next. I did find the “Surface Imaging of Proxima b and Other Exoplanets: Albedo Maps, Biosignatures, and Technosignatures,” on arXiv and here’s the link: https://arxiv.org/abs/1711.00185.
This gave me an idea of how life might have found another way to harvest the energy from red dwarfs in the far infrared and UV. Calcite crystals (crystalline aragonite) in sea shells may have developed properties that could transform the UV and infrared freqs into a useable forms.
Beyond Seashells: Bioinspired 2D Photonic and Photoelectronic Devices.
https://onlinelibrary.wiley.com/doi/abs/10.1002/adfm.201901460
Iridescent colors on seashells: an optical and structural investigation of Helcion pruinosus.
https://www.osapublishing.org/ao/abstract.cfm?uri=ao-41-4-717
Sea Shell Colours.
https://www.atoptics.co.uk/fz506.htm
Seeking structural color, engineers find it glinting in the sea.
https://www.seas.harvard.edu/news/2015/02/seeking-structural-color-engineers-find-it-glinting-sea
Encapsulated in the immortal words of Dr. Ian Malcolm: “Life finds a way”. We should always be cautious of using Earth as the universal model for life. It is an existence proof of life but probably does not encompass the possible space of biochemistry and form. Life evolves by modifying what it has to gain small advantages. This “bakes in” the molecular biology and biochemistry. Starting from a different set of working molecules, very different life forms could evolve.
Very interesting, thank you!
I feel confident that well before then publishers will have developed mechanisms to allow colorblind readers to easily distinguish between two graphs. They will look back on this era as a time of unnecessary roadblocks for some very talented people.
Otherwise it all seems reasonable.
Which particular graph or graphs posed a problem? Once upon a time when dinosaurs roamed the Earth, scientific papers were only published in monochrome. Different line types and shading were used to distinguish different series of data. I learned that even as late as the 1970s. Computer printouts created monochrome charts, and even early computer software created monochrome output. Generally, color output was advised to be simple, using extreme color variation to aid those with partial colorblindness. However, as we pack in ever more series to compare, this last tends to get ignored. I am not color blind, but I have difficulty determining which legend belongs to which line in a chart when many colors are used.
The charts use red/green to encode different variables. This is the most common colour blindness, IIRC. This website is useful:
http://colorbrewer2.org/#type=sequential&scheme=BuGn&n=3
I don’t deny the importance of this issue. Still, I do wonder if there’s a better way. What if you made a browser plug-in to automatically change problematic colors to something the users can recognize? It could have default settings for all the possible forms of color blindness. If a sufficiently clever algorithm went to work on a figure like this, maybe it could automatically recognize there are curves at different colors that can be distinguished by using just two; or maybe it could flip through various options until the user found something comprehensible … then share it online as an annotation for the website that other people with the same vision could automatically look up and use when they visit it. For monochromats or as a matter of personal preference, maybe the output could flash (at varying frequencies?) instead of displaying a color? It seems like it ought to work better to put the people affected in direct control rather than hoping everyone will do the right thing. Has no one in the world tried to do this?
Obligatory space reference: we’re all color blind in infrared to radio frequencies…..
There exist browser plug ins. Easy to find with any search engine. For example:
https://chrome.google.com/webstore/detail/colorblind-dalton-for-goo/afcafnelafcgjinkaeohkalmfececool?hl=en
Thanks – I had tried a search of extensions from within Firefox and didn’t see this, but per your prodding went back and found “Let’s get color blind” 1.0.0 which also seems to work, though reviews say it is not as good as the one which you recommended for Chrome. The Firefox plug-in has modes to simulate three types of color-blindness or to “daltonize” for them (to change the color output) or to do both. I just tried this (a one second download) – if it works, then a viewer with deuteranomaly should agree that Chl a and Bchl b are almost the same dark shade, and Chl b and lutein are the same light shade. This should definitely help me to review my own slide presentations going forward! It also has a mode to “simulate daltonization”, and if that works, then it appears possible to distinguish different shades at some setting of the slider, though never all at the same time. I was hoping for something a little more cutting edge AI, a little less 1.0.0, but at least it is something.
Excellent!
Looking at the top of the atmosphere spectrum of Earth vs Proxima b, the intensity in the near UV is roughly equal (100nm – 400nm). Could chlorophylls evolve to capture this part of the spectrum?
The problem with UV is that the energies are sufficient to break molecular bonds. Having said that, we know that insects see into teh UV range, so presumably, molecules that can detect UV and create a signal could also be used to capture the energy to fix carbon. I believe blue end of the spectrum that is absorbed by chlorophyll is responsible for the energy to split water to reduce carbon. So in answer to your question, while chlorophyll itself seems difficult to modify to absorb UV, another molecule may be able to do so.
It’s not a question of chlorophyll ( or related pigments) evolving to harvest UV so much as the raft of UV protective photopigments available to absorb and protect against its (very) harmful effects.
It should be said that many other ways of protecting against UV have evolved – involving amongst others, DNA regeneration – but the processes involved here are complex and not the subject of this post .
Carotenoid pigments for one offer protection against UV-A and early UV-B, but the big UV protective compounds used on Earth are the ’sinapate esters’. These have evolved to protect across UV-B to its ‘floor’ at 280nm. Given that it’s the most energetic form of UV to reach the surface of the Earth ( 5-7% incident sunlight) this is no surprise as there has been evolutionary pressure to find a solution as a result .Unlike NIR for photosynthesis in rare niche environments, no UV-C or beyond reaches the surface. So there has been no selection pressure to evolve Sinapoic acid or any other photopigment into the more energetic UV-C and beyond.
However, there has been at least one synthetic version created that operates below 280nm well into UV-C. It’s important to also note that in order to absorb such high energy wavelengths there is a price to pay . These esters are potentially genotoxic to their host life even on Earth. So whether any ‘sinapate plus’ would work in vivo remains a moot point for research . But in vitro is a promising start and it may be that these esters or analogues extending further into EUV represent a viable option on M dwarf planets. We shall see. Meantime they offer good effect in human sunscreens !
Certainly there are now many chemical and molecular engineering labs that specialise in photochemicals , as can be seen above with the fantastic work done creating synthetic bacterial chlorophylls that can operate into the near IR.
Mushrooms love UV light and other hard forms of radiation. I go out in the sun for a half hour and get a sunburn, but the majority of humans have enough dark pigment to not burn.
You worry about the lack of ozone in the atmosphere.
Would it plausible that life forms would evolve to produce ozone?
If the dominant life forms on Proxima b are chemosynthetic, we would be looking for different biosignatures.
If you are asking whether organisms could produce O3 for a local UV shield, I would say this is doubtful. Ozone is highly oxidizing. It destroys organic molecules. Even our own aerobic metabolism creates free oxygen radicals that must be mopped up to reduce cellular damage, but eventually it does damage our mitochondria as is one of the ageing effects. As soon as photosynthesis that releases O2 as a byproduct is evolved, the atmosphere will start producing protective ozone. However, it is easier for organisms to avoid UV by:
1. Hiding from it. Marine and aquatic life can stay at depths where the water column blocks UV. Terrestrial forms can live underground – e.g. worms, mole rats. Or be nocturnal (but more difficult on tidally-locked M-dwarf planets).
2. Evolve UV absorbing molecules. E.g. humans have melanin. Or evolve protection – shells, fur. However, eyes are always vulnerable to UV exposure. Perhaps animals in high UV environments evolve sonic senses instead.
3. Photosynthetic organisms might be able to create fluorescent molecules. These would both protect the plant and then emit visible light that could be used for photosynthesis. This is highly speculative.
I write about this in the CD post:
Detecting Early Life on Exoplanets
Hiding from UV is a protective mechanism that would work in theory . However in practice it would have limited utility – on an M dwarf planet anyway . Assuming early Cyanobacteria-like organisms kick life off – they would likely photosynthesise to live .
We’ve seen the potential here for the evolution of pigments able to extend photosynthesis into the infra red and chlorophyll variants could no doubt fill in the gaps to give complete spectral coverage from the bottom of visual to say 1100nm. Even then the low luminosity of M dwarfs is such that any putative alien photosynthesiser would need every last photon to eke out a living. Any protective shield would limit this and prove more of a disadvantage . ( though not on ozone free, UV bathed early Archean Earth- and modern day Mars? Anyway, here bacteria employed iron II salt rimes as effect shields ,with the necessary benefit of allowing through remaining plentiful visual spectrum light for photosynthesis ).
So that leaves some or all other techniques . Such as the DNA repair mechanisms seen in extremophiles populating nuclear reactors .
In terms of protective chemicals there are a good variety to choose. Beta carotenes and flavonoids mop up UV driven free radicals and ‘reactive oxygen species’ where present .Sinapate esters absorb UV as do a host of phenolic compounds with their central aromatic ring ( though with the unwanted side effect of genotoxicity – there’s nothing better than a benzene ring able to insert itself into a DNA helix and mess it up )
Finally tyrosinase enzymes are ubiquitous in the animal and plant world , playing a key role in the synthesis of melanin and related molecules which are as effective at absorbing UV in plants as in humans. ( but with the downside of making fruit go brown !)
That’s a lot of different chemicals which individually or better together could provide even a simple Cyanobacteria a formidable defence on an M dwarf planet . These chemicals have arisen through evolution on an Earth where just 5-7 % of incident sunlight is UV and even then mostly harmless UV-A. So imagine what could arise with election pressure from frequent EUV.
2 counter arguments:
Hiding should work very well. The key for photosynthesizers is to use differential filters. Water is one such ideal filter, blocking UV preferentially. As it blocks both UV and IR, photosynthetic pigments will absorb in the visible light, with organisms living at safe depths.
I think your reasoning is not correct. To be habitable, the effective solar flux is going to be high enough to warm teh planet with an appropriate atmosphere. The spectral temperature of Proxima is 3042K, which, if you use LED lights at home, looks slightly yellow compared to sunlight (and see image at top). The light intensity is not low. On Earth, photosynthesis can occur at a few percentage points of full sunlight. We see this near the bottom of the photic zone in the ocean, on the floors of tropical rainforests, and under ice floes. In all cases, the plants do not try to harvest more photons by absorbing more of the sunlight’s spectrum. Always bear in mind that evolution is driven by the selection of individuals to do slightly better than other members of its own species as well as other species. If photosynthesis is lower around M-dwarfs, it won’t matter as all plants will be similarly affected. On Earth, photosynthesis is very inefficient, yet we do not see plants trying to compete by increasing photosynthetic efficiency (except with C4 vs C3) or increasing spectral absorption.
As with all speculation, we are likely to be only partially correct and will be surprised by reality. I look forward to the data that will start to resolve these issues. I expect this to come from telescopes, using a variety of techniques, which will ultimately be confirmed and details added with probes to living worlds. Sadly, I won’t live long enough to see the results of any possible interstellar probe. I will have to live with the hints we get from remote sensing.
Thanks Alex: I’m sure we will both live to see E-ELT /Metis driven spectroscopy of Proxima, Ross 128 , Lalande et al with sufficient resolution to spot a photosynthetic ‘edge’ if one exists.
I take your argument about intensity and planetary temperature . However as a caveat to this , the longer IR wavelengths emitted as a majority part of the stellar SED of all but the earliest M dwarfs – the more so the later the spectral class – are absorbed more by terrestrial style atmospheres . They also have a much smaller ‘ice albedo’ than shorter wavelengths to increase surface absorption too . So in terms of heating a little energy goes a lot further. More than for photosynthesis certainly.
Any physical screen – be it dust, salt rime/rust – and water too , is unavoidably going to absorb some incident light, including the shorter more energetic photosynthetic efficient variants . With a luminosity less than a fifth of a percent of Sol – and 1/200000 in terms of visual light – that doesn’t allow for any of the attrition seen on Earth, which just hasn’t seen the same kind of stringent evolutionary pressures given Sol’s generous emission centred around visual wavelengths.
I’m postulating that physical screening does play a big UV protective role – but for more primitive chemotropic life in the first instance . At some point, a tiny fraction of this life gradually loses some of its physical shield ( as with atmospheric Cyanobacteria for Archean Earth) thus increasingly exposing it to EUV . However, having developed EUV absorbing pigments as an accidental offshoot of the chemosynthesis process, some of it is able to survive long enough to make EUV an evolutionary driver. Unlike on Earth which has never seen anything worse than UV-B and even then only in small amounts . But even this limited UV exposure helped create a huge range of protective strategies. More of them chemical than physical too. Goodness knows what UVB could lead to. ( and what chemists have achieved in the lab with
bacteriochlorins !)
Finally, after all this, chlorophyll or some analogue appears as on Earth to start photosynthesis and harvest the ubiquitous surface energy supply. A poor cousin compared to Earth as you say, but conferring just as big or indeed more of an evolutionary advantage given it will likely have to evolve to be far more efficient .
I grant that physical shields could play a role in early M dwarfs nearer to 4000 K, which emit a much greater fraction of their SED in the visual with luminosities around a tenth that of Earth.
Looking for recent publications I stumbled across this one (obscure journal though): https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6042481/ They say they improved UV resistance 50-fold in a bacteriophage, which certainly does not have much thickness to work with. (They talk about mutations but I don’t see them mention thymine dimers) To be sure, the paper about E. coli from 2011 was less inspiring: https://onlinelibrary.wiley.com/doi/full/10.1111/j.1558-5646.2011.01438.x With recent advances in artificial base-pairs in DNA, I think it is very plausible that alien organisms could find an alternative to pyrimidines. Even eyes can be given sun protection – our own lenses come with 3-hydroxykynurenine to block the UV, which is why patients with cataract surgery had to be recruited to view the secret UV signals during World War II.
Well some good news for speeding up process for screening the planet candidates around M and K dwarfs suns. This will give us a good idea as to what ranges the atmospheres are depleted around the inner edge of the habitable zone.
Identifying Candidate Atmospheres on Rocky M dwarf Planets via Eclipse Photometry.
https://arxiv.org/abs/1907.13138
Novel Method Proposed To Find Atmospheres On Rocky Worlds.
http://astrobiology.com/2019/12/novel-method-proposed-to-find-atmospheres-on-rocky-worlds.html
In many previous comments on this website, I have mentioned that adding a CURRENTLY FUNCTIONAL high amplitude pupil mask to the ZIMPOL polarimiter on the VLT SPHERE/ZIMPOL instrument package could realistically obtain the first ever ACTUAL image of Proxima b in POLARIZED LIGHT! NOW: Enter arxiv: 1912.01232 “PlanetEvidence: Planet or Noise?” by Jacob Golum et al. this leads me to believe that this may be a WHOLE LOT MORE REALISTIC than I previously thought. OH, BY THE WAY: The CURRENT VLT SPHERE/ZIMPOL setup recently observed both Alpha Centauri A and Alpha Centauri B in an(unfortunately failed)attempt to “scoop” NEAR, whose observation campaign(back in May)results have quite mysteriously not yet(they were supposed to be published by October)been made public.
Thanks for the terrific review of photosynthesis Alex. I had forgotten most of my plant physiology so this has helped a lot. Can’t wait for the first information about the atmosphere of the nearest exoplanet that we know of. I wish we were closer to the time of launching the Starshot probes.
https://arxiv.org/abs/1912.01783
Searching for a standard Drake equation
Jose Antonio Molina Molina
(Submitted on 2 Dec 2019)
In the 20th century the scientific search for extraterrestrial intelligence began, and the Drake equation was proposed to estimate the number of extraterrestrial species humanity could attempt to detect, N. This paper discusses the need to update and standardize this equation.
A new and more accurate expression is deduced which contains the classic equation as a particular case, and its advantages are discussed. A necessary condition is also stated for its use in scientific contexts: if N is defined as the total number of civilizations like ours then N = 1, given that we exist, and consequently the working hypothesis of the SETI project can be expressed as N>1.
In this case, the Drake equation is being applied in a scientific way, because it is trying to test a hypothesis based on evidence.
Comments: 15 pages, 3 tables
Subjects: Popular Physics (physics.pop-ph); Earth and Planetary Astrophysics (astro-ph.EP)
Cite as: arXiv:1912.01783 [physics.pop-ph]
(or arXiv:1912.01783v1 [physics.pop-ph] for this version)
Submission history
From: José Antonio Molina [view email]
[v1] Mon, 2 Dec 2019 19:15:47 UTC (476 KB)
https://arxiv.org/ftp/arxiv/papers/1912/1912.01783.pdf
His value fb is interesting. The chain requires assuming more than one biosphere exists (reasonable), that there can be other than carbon biospheres (interesting in itself), and then the need for a fraction of surface, carbon-based biospheres of intelligent life.
While I doubt that Clarke’s short story about lithospheric intelligent species could exist, and the assumption that carbon-based life is a requirement for a civilization, both these constraints may be false. In particular, it assumes, as I think Drake did (still does?) that civilizations are confined to their homeworlds, or at least their home systems. That we are even thinking about probes to other stars and the very purpose of interstellar groups is about human expansion to the stars, this seems an obsolete, unwarranted assumption. If machine civilizations exist, then most of the factors in any of these equations are just unneeded, with planets unneeded for bases other than as resources. Removing these constraints removes most of the factors in the equations (Drake’s and this new one). While I don’t believe Jim Benford’s suggestion of looking for “lurkers” in nearby asteroids will find anything, at least it ignores that assumption that ETI is only located around another star, and not potentially distributed out in space, exploring the galaxy as we are intending to do with flybys and eventually orbiting and landers on exoplanets.
His conclusion that SETI is trying to prove N>1 is correct, regardless of the equation used to estimate intelligent civilizations in the galaxy.
The original Drake Equation held to the paradigm that restricted SETI for decades, namely that they were searching for and calculating estimates based on versions of us.
Supposedly a group of altruistic alien intelligences who also conduct science the way it is done in the West would be beaming messages of introduction plus tons of their knowledge all over the galaxy, because, hey, isn’t that what all scientists and like-minded engineers do, irregardless of their place of origin and upbringing?
Even though humans, by contrast, have only done METI a relatively few times and sporadically at that. Then everyone else flips out about how marauding aliens will come and enslave or eat us, even though we’ve been making ourselves known to the Milky Way for over a century now technologically. Smart aliens would also know there is at least life on Earth going much further back, none of which has been under our control.
It is definitely time to upgrade the paradigm. And oh yeah, give it some serious funding and let more than just a group of radio astronomers join the fun. Some day we may wish we had done a lot more in the SETI effort.
Scientists Are Contemplating a 1,000-Year Space Mission to Save Humanity
Relocating the human race to a more hospitable planet would mean that multiple generations would be born in-transit.
by Corin Faife
December 5, 2019
https://onezero.medium.com/scientists-are-contemplating-a-1-000-year-space-mission-to-save-humanity-70882a0d6e47
To quote:
Others looked at what it means for the crew of the ship — not the first generation, who choose to leave Earth behind, but for the second, tenth, fiftieth, one hundredth, the people for whom our planet is just a myth; for whom there will be no other life but the journey.
Andrew McKenzie and Jeffrey Punske, linguists from the University of Kansas and the University of Southern Illinois, write that “[i]f a trip takes several generations to complete, the language may differ significantly at arrival from that of the passengers at departure.”
More evocatively they suggest: “Even if the onboard schools rigorously maintained the teaching of ‘Earth English’ the children would develop their own Vessel English dialect, which would diverge from Earth English over time.” The problem would be compounded by the fact that this “Vessel English” — using English as just one example — would be unique to each ship, so that the crew of two ships arriving at the same planet would speak a different dialect, or even a different language altogether.
Ultimately, to survive we will need to relocate.
Separately, Neil Levy, a professor of philosophy at Macquarie University in Sydney and senior research fellow in ethics at Oxford University, considered the moral implications in an article for Aeon:
“A generation ship can work only if most of the children born aboard can be trained to become the next generation of crew,” he writes. “They will have little or no choice over what kind of project they pursue. At best, they will have a range of shipboard careers to choose between: chef, gardener, engineer, pilot, and so on.”
In other words, their life options will be extremely limited, as would be the range of experiences they can enjoy. Would it even be ethical to put them in this situation?
The conclusion depends on what we believe is justified to preserve our species, a reckoning Levy declines to make. Instead, he points to the subtext of the question: Life outcomes are already defined by accident of birth in the world as it is; the range of any child’s possible futures is constrained by poverty, nationality, religion, culture. This may be unjust, but we accept this as part of the human condition. “Asking about the permissibility of generation ships,” he writes, “might give us a fresh perspective on the permissibility of the constraints we impose now on human lives, here on the biggest generation ship of them all — our planet.”
Generation ships always struck me as absurd. If you can live and survive in space for a thousand years with zero access to any outside resources, why would you need to go to a planet at all? Why leave the outer Solar system? Naturally that’s a big if: imagine the President of Upper Right Quadrant promising to work with his fellow nations to crack down on the illegal scrap mining operators and reduce air leakage by 25% within the next 15 years. Nothing’s arriving at the far end but skeletons and manifestoes.
By contrast, even a post-K/T Earth offers an environment that can (at least) be readily terraformed, has air that is breathable with minimal processing (1), abundant water and mineral resources. Natural and artificial tunnels provide ample room for large colonies protected from any continuing bombardment (Strangelove, 1964). You may have to cull some hostile native fauna, but such is par for the course for colonization projects (the generation ship isn’t planning to turn around when they find lifeforms, are they?)
1. https://www.theguardian.com/australia-news/2019/dec/08/just-leave-delhi-beijing-and-mexico-city-residents-on-how-to-cope-with-pollution
The need for generation ships is based on 2 constraints:
1. That the energy to reach near lightspeed to gain the needed time dilation effect is not practical based on any energy source plus needed engineering.
2. Without real or perceived life extension, humans will live and die on all but the shortest voyages.
Constraint 2 can be mitigated by:
a. life extension technology
b. cryosleep
c. embryo/seed ships
d. mind transplanting into machine bodies
Some of the constraint 2 mitigation also removes the requirement for the expected large size of generation ships, e.g. seed ships might be quite small. Encoding minds for storage would also remove much of teh mass requirement. The fewer resource-consuming entities on board the ship, the fewer the constraints on recycling are needed.
For those of us prepared to accept that machine intelligences are going to be the true star farers, the easier it is to contemplate slow, small, ships traveling to the stars and potentially colonizing space.
The need to leave the solar system is going to be a requirement if populations want to avoid eventual resource limitations and hegemonic polities. This is the same desire of the Mars Society people to escape the constraints of Earth societies and “start again”, which in turn has historical roots, such as the Mayflower voyage for the initial founding of the New England colonies. I don’t expect these long-established historical precedents to disappear in our future. O’Neill saw the establishment of his space colony habitats as the basis for eventually leaving our solar system, as a generation ship is just a colony with a stellar flight capable propulsion system.
As the saying goes, “If it can be done, it will be done”.
I would like to know if all these folks ever read or at least made themselves aware of all the science fiction works on multigenerational starships, starting with Heinlein’s classic Orphans of the Sky.
For this is what science fiction does best, playing out various scenarios without actually experimenting on living subjects.
https://www.kirkusreviews.com/features/generation-starships-fiction-and-fact/
https://sciencefictionruminations.com/sci-fi-article-index/list-of-generation-ship-novels-and-short-stories/
http://www.sf-encyclopedia.com/entry/generation_starships
That really depends on the size of the ship. Obviously a ship the size of a planet will not require that constraint – the Earth is an example of that. Given teh skillset needed in teh population on arrival, most likely most passengers will not need to become trained other than to select skillsets needed to pass on to the next generation if needed. But really good education systems might just avoid that requirement.
Looking for exoplanet life in all the right spectra
By Blaine Friedlander | December 4, 2019
A Cornell senior has come up with a way to discern life on exoplanets loitering in other cosmic neighborhoods: a spectral field guide.
Zifan Lin ’20 has developed high-resolution spectral models and scenarios for two exoplanets that may harbor life: Proxima b, in the habitable zone of our nearest neighbor Proxima Centauri; and Trappist-1e, one of three possible Earth-like exoplanet candidates in the Trappist-1 system.
The paper, co-authored with Lisa Kaltenegger, associate professor of astronomy and director of Cornell’s Carl Sagan Institute, published Nov 18 in Monthly Notices of the Royal Astronomical Society.
“In order to investigate whether there are signs of life on other worlds, it is very important to understand signs of life that show in a planet’s light fingerprint,” Lin said. “Life on exoplanets can produce a characteristic combination of molecules in its atmosphere – and those become telltale signs in the spectra of such planets.
“In the near future we will be seeing the atmosphere of these worlds with new, sophisticated ground-based telescopes, which will allow us to explore the exoplanet’s climate and might spot its biota,” he said.
Full article here:
http://news.cornell.edu/stories/2019/12/looking-exoplanet-life-all-right-spectra
Thanks Alex
A very interesting read along with the comments too
Thanks
How quantum dots supercharge farming, medicine and solar, too.
Magenta is not a real color but the absence of green leaving the opposite ends of the visual spectrum red and violet light. We see this as magenta and plants thrive under it’s illumination.
https://www.cnet.com/news/how-quantum-dots-supercharge-farming-medicine-and-solar-too/