The right kind of atmosphere may keep a planet habitable even if it crowds the inner region of the habitable zone. But atmospheric evolution involves many things, including the kind of geological activity our own planet has experienced, leading to sudden, deep extinctions. Centauri Dreams regular Alex Tolley today takes a look at a new paper that examines the terrestrial extinction of marine species in the Permian event some 252 million years ago. As we examine exoplanet habitability, it will be good to keep the factors driving such extinctions in mind. Tolley is a lecturer in biology at the University of California and author, with Brian McConnell, of A Design for a Reusable Water-Based Spacecraft Known as the Spacecoach (Springer, 2016). A key question in his essay today: Is our definition of the habitable zone simply too broad?
by Alex Tolley

In the search for life on exoplanets, questions about whether the planet is within the HZ given a plausible atmosphere is based on timescales as a fraction of stellar lifetime on the main sequence. With water may come the emergence of life as we know it, and then the long, slow evolution to multicellular life and possible technological civilization. Planets may initially form too close to a pre-main sequence star to be in the HZ, then enter the HZ, only to leave it again as the star increases in luminosity with age. Earth has experienced about a 30% increase in solar luminosity over its lifetime. The CO2 level needed to maintain a constant surface temperature via the greenhouse effect has had to decline to offset the increased insolation. In 1 to 2 billion years, the further increase in solar luminosity will require the CO2 levels to decline below that needed for photosynthesis, or the Earth’s surface will heat up beyond that sustainable for life.
Yet when considering the environment on a world in the HZ, we should be cognizant that climatic instability may create shocks in the short term that have major impacts on life. Earth has experienced 5 major extinctions based on our reading of the fossil record. The most famous being the dinosaur-killing KT event that ended the Cretaceous and allowed mammals to evolve into the newly vacated ecological niches. However, the largest extinction is known as the Permian extinction, or “Great Dying” when over 95% of marine species became extinct about 252 mya. Unlike the KT event, which was a cosmic throw of the dice, the Permian extinction is believed to be due to massive volcanism of the Siberian Traps that released vast quantities of CO2 into the atmosphere that increased its concentration at least several fold. This caused a rapid temperature rise of 10s of degrees Fahrenheit and was accompanied by ocean acidification.
A new paper by Julian Penn et al suggests that this global temperature change caused the extinction of marine species primarily by metabolic stress and hypoxia.
The core idea is that multicellular, aerobic organisms require critical oxygen pressures to live, with their lowest levels of metabolism during resting, and higher levels for activities, such as swimming or feeding. Sessile organisms may have just a 1.5x increase in active metabolic rate over resting, while energetic organisms like fish may be 5x or more. As temperatures rise, so does the metabolic rate. This, in turn, requires adequate oxygen for respiration. But as the temperatures rise, the dissolved oxygen levels fall, placing additional stress on the animals to maintain their respiration rate. Penn integrated climate models to compute the temperature change and dissolved oxygen partial pressures, with the estimated metabolic rates for the activity of various modern animals to represent Permian species, to determine how ocean habitat temperatures impact the metabolisms of marine genera and probable extinction rates.
Figure 1 shows the relation between metabolic rate and temperature, and the temperature increased metabolic index of ocean habitat by latitude and depth. The polar latitudes and shallower depths show the highest changes in the metabolic index, indicating the most stressed habitats.
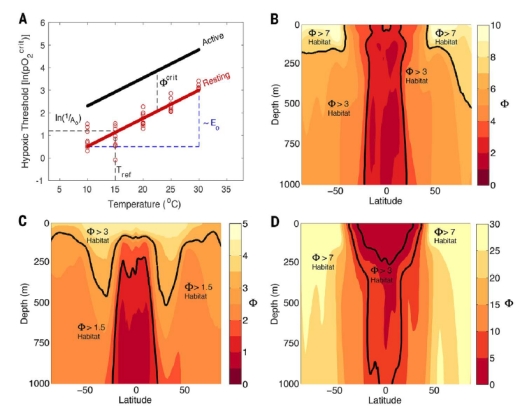
Figure 1. Physiological and ecological traits of the Metabolic Index (F) and its end-Permian distribution. (A) The critical O2 pressure (pO2 crit) needed to sustain resting metabolic rates in laboratory experiments (red circles, Cancer irroratus) vary with temperature with a slope proportional to Eo from a value of 1/Ao at a reference temperature (Tref), as estimated by linear regression when F = 1 (19). Energetic demands for ecological activity increase hypoxic thresholds by a factor Fcrit above the resting state, a value estimated from the Metabolic Index at a species’ observed habitat range limit. (B) Zonal mean distribution of F in the Permian simulation for ecophysiotypes with average 1/Ao and Eo (~4.5 kPa and 0.4 eV, respectively). (C and D) Variations in F for an ecophysiotype with weak (C) and strong (D) temperature sensitivities (Eo = 0 eV and 1.0 eV, respectively), both with 1/Ao ~ 4.5 kPa. Example values of Fcrit (black lines) outline different distributions of available aerobic habitat for a given combination of 1/Ao and Eo. Credit: Justin Penn and Curtis Deutsch, University of Washington.
Figure 2 shows the spatial changes in ocean temperature and oxygen concentrations. Oceanic temperatures rise, particularly towards the poles, and with it a reduction in dissolved oxygen. As expected the greatest rises in temperature are at the shallower depths, particularly with the highly productive continental shelves. Oxygen level declines are most widely seen at all depths at the poles, but far less so in the tropics.
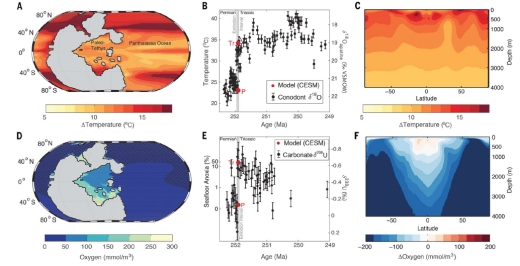
Figure 2. Permian/Triassic ocean temperature and O2. (A) Map of near surface (0 to 70 m) ocean warming across the Permian/Triassic (P/Tr) transition simulated in the Community Earth System Model. The region in gray represents the supercontinent Pangaea. (B) Simulated near surface ocean temperatures (red circles) in the eastern Paleo-Tethys (5°S to 20°N) and reconstructed from conodont d18Oapatite measurements (black circles) (4). The time scale of the d18Oapatite data (circles) has been shifted by 700,000 years to align it with d18Oapatite calibrated by U-Pb zircon dates (open triangles) (1), which also define the extinction interval (gray band). Error bars are 1°C. (C) Simulated zonal mean ocean warming (°C) across the P/Tr transition. (D) Map of seafloor oxygen levels in the Triassic simulation. Hatching indicates anoxic regions (O2 < 5 mmol/m3). (E) Simulated seafloor anoxic fraction ƒanox (red circles). Simulated values are used to drive a published one-box ocean model of the ocean’s uranium cycle (8) and are compared to d238U isotope measurements of marine carbonates formed in the Paleo-Tethys (black circles). Error bars are 0.1‰. (F) Same as in (C) but for simulated changes in O2 concentrations (mmol/m3). Credit: Justin Penn and Curtis Deutsch, University of Washington.
The authors conclude:
The correspondence between the simulated and observed geographic patterns of selectivity strongly implicates aerobic habitat loss, driven by rapid warming, as a main proximate cause of the end-Permian extinction.
However, while the temperature is the proximate cause, the authors note that other factors are also involved.
“In our simulations, net primary productivity is reduced by ~40% globally, with strongest declines in the low latitudes, where essential nutrient supply to phytoplankton is most curtailed.”
Ocean acidification is also a potential factor, as we may be seeing today. Acidification will be higher at the poles, creating a habitat barrier for species that require more calcification.
Figure 3 is a schematic of the model, fitting the probable extinction rates to the fossil record. Their model predicts a latitudinal impact of warming that is also suggested by the fossil record. Their explanation for this spatial pattern is that tropical organisms are preadapted to warmer temperatures and lower O2 levels. As the oceans warm, these organisms migrate polewards to cooler waters. However, polar species have nowhere to migrate to, and therefore have a higher rate of extinction.
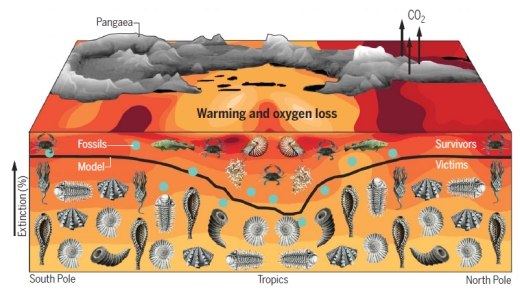
Figure 3. An illustration depicting the percentage of marine animals that went extinct at the end of the Permian era by latitude, from the model (black line) and from the fossil record (blue dots). The color of the water shows the temperature change, with red representing the most severe warming and yellow less warming. At the top is the supercontinent Pangaea, with massive volcanic eruptions emitting carbon dioxide. The images below the line represent some of the 96 percent of marine species that died during the event. Credit: Justin Penn and Curtis Deutsch, University of Washington.
As our current analog of the Permian climate change impacts the oceans, we are already seeing warm water species appearing in the cold North Atlantic, far north of their historic ranges. We can also expect species like the Arctic ice fish that has no red blood cells due to the high O2 concentrations in polar waters to become extinct as polar waters continue to warm.
What about the extinction of terrestrial life? 70% of terrestrial faunal species went extinct. The attractiveness of this theory is that it also applies to terrestrial life, although the oxygen depletion was not a factor. What is clear as well is that the CO2 increase heated the planet, overwhelming any cooling from dust blown up into the atmosphere, as experienced with the 2 year global cooling after Mt. Pinatubo erupted.
Had the Earth been closer to our sun, or temperatures risen further due to greater volcanic activity, the extinctions might conceivably have been 100% for all multicellular genera. Earth life might have been pushed back to primarily archaea and bacteria. The atmosphere might have reverted back to its Archaean state. If photosynthesizers were still present, how long would it take for aerobic multicellular life to evolve again?
The major extinctions have implications for life on exoplanets. Worlds closer to the inner edge of the HZ may be supportive of life if the atmosphere stays stable. However, as we have seen with the example of the Permian extinction, geologic processes can upset that balance, potentially making a world uninhabitable for a period, forcing any life to be restricted to simpler forms. How frequently could such events cause mass, even total extinctions, on other worlds, despite long-term conditions being favorable for life? It is perhaps worth considering whether the inner edge HZ limits should be made more conservative to allow for such events.
The paper is Penn et al., “Temperature-dependent hypoxia explains biogeography and severity of end-Permian marine mass extinction” Science Vol. 362, Issue 6419 (7 December 2018). Abstract (Full Text behind paywall).



Another well reasoned article that adds to the evidence for the “rare Earth” theory as a likely answer to the Fermi paradox.
Agree with you Joe. And to keep this rare gem of a planet near the sweet spot of the HZ for as long as possible the Earth’s orbit will need to be expanded. We already know how, just rob momentum from Jupiter by means of a third body on a correctly engineered orbit.
@Bruce D. Mayfield ,
I would suspect that long long before we ever get to the point of being able to induce gravity assisted orbital changes to our planet, the human race will have gone extinct. That’s not being a Debbie Downer by the way; my suspicions is that, with regards to organisms in general, just as life can arise, death can arise probably just that much easier, and may be due to entropy (if you allow me to call it that). In the case of humanity, we have inadvertently I feel contaminated the environment so thoroughly that now animal species and humans are having a difficult time reproducing themselves. In a sense, there is a racial exhaustion that is occurring among the flora and fauna of this planet. And I suspect it’s not going to end well. I came across an article recently in the magazine Scientific American, which explored the possibility that planets as a whole become in their lifetime ‘old’ just as organisms do and they began to lack a certain vitality because their , geological mechanisms begin to wind down. I found it extremely insightful because I don’t think most people have ever considered the possibility that planet, just like organisms can get old also. Just a thought here.
We humans certainly are trending toward disaster Charley, so your reply is reasonable. Your right, the current man made mass extinction event will (allmost) wipe us out too. But ultimately, I’m an optimistic sort. Survivors will have learned lessons, pieces will be picked up, life will go on, and eventually flourish. Our wiser descendants will see how precious life is, and they would be the ones who would take the actions needed to preserve Earth’s habitability.
Otherwise, your right, we’re ultimately doomed. But nah, our potential is so much better! We can ‘go forth and fill the galaxy and subdue it.’ Andromeda too for that matter, because it will start merging with the Milky Way before the Sun exits the MS.
Ya gotta look on the bright side Charley!
Life may well populate the Andromeda galaxy, but it will not be human as we know it. Species don’t last that long, and humans are likely to evolve into new forms well before then. Our lineage may even go extinct, although I would hope that by finding new places in our galaxy to colonize, that may never happen.
I agree that this article well reasoned, but it is about our own planet, conditions in the outher star system will be different so we cannot apply our particular case to other worlds.
To apply this article to all possible ET planets in all systems we have to be sure that Solar System developmnet is repeated same steps and ways multiple time in the Universe, meanwhile – science does not have any evidence and proves to this statement. Most probable everything everywhere is developping using the same laws but different ways.
So we canot apply our case to everything, opposite , if finally we apply the Erth scenario to every ster system, we how to conclude – the Earth type life is everywhere – that contradicts Fermi Paradox :-)
Yes, the article is only about Earth, but IMO the general theme of the article can be applied universe wide. Planets change over time. The stars they orbit change over time. Too much change or too fast change tends to be bad for life. This means the duration of a habitability zone in time is probably less than previously thought.
Only one example – is bad reference to deduce universal laws.
Foe example I can suppose that possible variant where duration of HZ can be longer than on the Earth, by the way on the Earth HZ has not finished yet :-) So your conclusion is not so valid too…
We may need a “Fermi Award” (a more severe version of the Darwin Award).
Let’s not win it!
Michael
A wonderful post. But like I said, there may be only three of four civilizations in this galaxy or in any galaxy and if they’re sub-light spacefarers we won’t hear about them for many centuries. Of course lightning can strike, which is why we hope we’ll be lucky such ftl creatures appreciate mammals and are friendly too. There are trillions of galaxies. And trillions of civilizations out there. We just haven’t been lucky yet. A little cosmic patience will help one endure the silence.
Perhaps, we haven’t been unlucky yet?
Some of these questions may be answered by building Biosphere 2 type habitats in space. That should be a major project begining in this century.
Agreed. Closed loop life support is going to be critical for deep space exploration.
May be Mars’s life support reformation?
Long standing bases on the Moon?
Small self contained hollow earth type habitats orbiting independently of planets. The need is to experiment with life in contexts we can control.
Yes in orbit around Earth or the Moon make most sense, if there’s an emergency the personnel can then be more easily evacuated.
How to sell the idea to a government organization or corporation?
To state it be the first space colony, it need something more to sell the idea – perhaps it could double as one solar energy platform. That could get a person like Elon Musk or any of his competitors interested.
Failing government help maybe a multi billionaire would be interested.
I had in mind starting with fungi, moss, insects, and arachnids. A spin would be necessary to simulate gravity and separate land from water while windows or sunlamps could provide night and day.
Using simple forms of life would enable us to get a handle on things without endangering more complex creatures. (We could also use plants and insects we might raise for food and sell.)
This is likely a centuries long project.
I could design such an ecology as you describe with smaller critters, and might even get it to work.
What’s necessary is the size for the experiment compared to the organism involved, because just like in nature, there will be population swings.
If the container is too small, the herbivores would eat up all available food when they get more numerous, and then starve to death.
While in nature they move on to new areas, while the other area recover.
Extreme example: the African wandering grasshopper Locusta migratoria, or the common Lemming Lemmus lemmus.
The best test would not be with moss and fungi as those grow quite slowly, to get a result in a better time – it should have more fast growing plants. Then again, we never get away from the fungi, those will be vital for breaking down end products that otherwise would build up.
I don’t have sufficient engineering knowledge to design the habitat in any detail so I’ll defer to those who do.
You could design the habitat to be in separate sections with only intermittent access between them, to mimic geographic separation of the populations, yet offer some leakage in teh event of a section having a permanent population crash or epidemic.
I don’t agree with the rare Earth theory. Even if we include the necessity of a Moon for a magnetic field and other limitations like the right size of the like G class, and HZ there still would be one million Earth twins in our galaxy alone which is just an educated guess with a little probability.
It would be funny if there was intelligent life out there and they all thought that there was nothing out there and did nothing.
Your certainly not alone in that. “Rare Earth” was and is quite controversial. But it can be somewhat hard to look at all the data completely objectively. We humans tend to believe what we hope to be true and we tend to downplay or even ignore evidence to the contrary.
I hope there are one million or even more Earthlike planets in this galaxy! That would be awesome, but that’s not a scientific reason to expect that it must be so, as I’m sure you’d agree.
Or as some cultures use to think, that it was all created for them, ” The Crown of Creation” which is all the “Rare Earth” amounts too. We are the first and only species on this Earth that are capable of stopping a mass extinction from asteroids or comets. We are also very capable of doing the same to earth with our weapons. So we are no different then the worshipped gods of old as Oppenhimer said ” The destroyer of worlds”.
I’ve been mulling over a reply to this for days Michael. This statement sticks in my craw: “… ‘The Crown of Creation’ which is all the ‘Rare Earth’ amounts too.” That’s dismissive and very inaccurate. Wikipedia gives “Rare Earth hypothesis” a much more balanced treatment;
“In planetary astronomy and astrobiology, the Rare Earth hypothesis argues that the origin of life and the evolution of biological complexity such as sexually reproducing, multicellular organisms on Earth (and, subsequently, human intelligence) required an improbable combination of astrophysical and geological events and circumstances.
According to the hypothesis, complex extraterrestrial life is an improbable phenomenon and likely to be rare. The term “Rare Earth” originates from Rare Earth: Why Complex Life Is Uncommon in the Universe (2000), a book by Peter Ward, a geologist and paleontologist, and Donald E. Brownlee, an astronomer and astrobiologist, both faculty members at the University of Washington.”
The article https://en.m.wikipedia.org/wiki/Rare_Earth_hypothesis is a good read, fairly outlining both pros and cons.
Let’s put it this way the only places that we have actually searched for life is Mars and we have landed on Venus and Titan. The gas giant planets could have life but no long term probes have been sent to investigate. Life in interior oceans of numerous satellites and comets and asteroids but no sample returns yet. Sounds to much like the old dogma just reasserting itself. When it comes to Complex Life we have barely scratched the surface to put it mildly, and if panspermia is common then life on the gas giants is common also! Do not get me wrong, the earth is Unique and global warming is a real threat, but just the idea that a crazy, conceded naked ape is it in the universe is way to far from the truth.
You cannot make that assumption. Life may have reached Jupiter but found it inhospitable. It would certainly be worth looking for, especially in the atmosphere where H2O clouds can form. While we find bacteria in the atmosphere of Earth, we don’t know if they replicate there, or are just temporary visitors from the surface, the latter cannot be the case on Jupiter.
Cannot accept your point of view.
– I am sure the rare Earth – it is fact. Exactly like the case that two similar fingerprints are not existing, same case with two similar planets.
– In same time wandering, why should someone conclude that “rare Earth” and “ET life” are antagonists?
Our Earth’s case tell as about our Earth’s case, and tell nothing about alternative variants.
We can’t conclude anything definite about ET life until we find it.
There seems to be none in our system and we ought to be thankful because we can modify nearby worlds to suit ourselves without killing alien life.
I do not conclude anything about ET life, my notes are not related to existence of absence of ET life, only want to tell that single , our Earth life evolution scenario cannot be applied in same pattern to other worlds, for sure they be different.
This issue has no any connection to the ET life existense or not existense, in connection to life – I agree with you that this fact can be proved only by real finding ET life, not by speculation on the paper and inventing pseudoscience like astrobiology …
I agree with this idea since it is not hard to throw the climate out of balance with a warm Earth. A cold Earth might remain in an arrested state of evolutionary development.
A thoughtful review of significant issues; thank you, Alex T!
Some posit that carbon dioxide will be scrubbed from the atmosphere by geologic processes, eliminating photosynthesis-dependent life on this planet:
The great chemical reaction: life and death of Gaia
https://cassandralegacy.blogspot.com/2012/05/great-chemical-reaction-life-and-death.html
while others posit other mechanisms for the extinction of life on this planet:
Causes and timing of future biosphere extinction
https://www.biogeosciences-discuss.net/2/1665/2005/bgd-2-1665-2005.pdf
And even if our species continues indefinitely through astronomical space and time, genetic drift and spatial separation will result in speciation, perhaps as behaviorally different as the lion and the lamb.
Man after Man
https://en.m.wikipedia.org/wiki/Man_After_Man#/search
The “man after man” idea is fun, but this is not very probable as humans will unavoidably merge with technology and already has stopped the natural evolution with modern food, healthcare asf. Natural evolution of man is outrun by far already. How humans are build today is the end, there will be no more. In hundred years the homo sapiens sapiens is something else, and it will not be an evolutionary change, but a man made change.
Don’t you suppose that “man maid change” also one of pathways of “natural evolution”?
Opposite you claiming – human race is something unnatural – that I cannot accept, I suppose it is religion’s claim, not scientific.
Contrary to popular opinion (and what I was taught back in the 1970s), evolution has not stopped on Earth. . I think what people mean is that selection has been reduced. But recall evolution requires 3 features:
Natural variation
Inheritance of traits
Selection allowing for differential reproductive survival.
Natural variation has increased. Inheritance has not changed (yet). Selection is what people think of as having declined. This may be so, but there are other forms of selection than competition – sex selection, and now, arguably, economic selection. (The wealthy are far more likely to reproduce if global conditions get worse.)
I almost forgot – random drift. This is particularly important for small populations, especially those that have been isolated by some event. Drift may/will result in speciation from the parent population over time. It has been hypothesized that the Northern Plains American Indians features are due to genetic drift starting from a small founder population. Selection, by whatever means, drives evolution as individuals in a population compete with each other for limited resources in a generally Malthusian world.
Yesterday on CBS they reported that Americans are .01% shorter, .01% living less longer, and are 30 pounds fatter than in the 1960s. Is this trending toward ‘evolution’?
Your student.
Those are probably the result of nutrition and the effects of immigrant populations. The opioid crisis is shortening the lifespan of non-Hispanic white males.
Oh yes, genetic drift is still around.
So lets agree that also humans still got microevolution at work.
Yet with people relocating and migrating at an even faster pace, combined with the fact that the migrating people more readily will find a partner, even if they would have been seen as less desirable, considered beautiful etc in a more rural/closed society. The selection mechanism might stop working about in our time.
Selection for economic success, or other traits will still occur. Yes, large populations are more stable, but that doesn’t stop self-imposed isolation – e.g. physical or social enclaves with social strata. An extreme case is teh royal intermarriages, although that selection did not in any way seriously change the relevant population. More likely population changing effects might be selection for beliefs that requires underlying brain architecture selection.
Evolution in humans has become faster:
Recent acceleration of human adaptive evolution
“World Population Might Go to 40 Billion by 2150 But We Can Already Feed All Those People”
I see several parts to the problem where many people believe the doomer overpopulation myths.
The fear story resonates with deep caveman fears of starvation. Any individual person can experience hunger. This reinforces and validates the fear of potential starvation.
The small scale and valid fears and issues for individuals and small groups do not match up to real global risk. There are counter-intuitive aspects.
Is an objection – technical or some other subject relevant? I disregarded areas that I do not impact the core question of whether overpopulation is a true risk.
Scale of overpopulation issue for the world. 30% increase by 2050, 60%-200% increase by 2100. 100%-400% by 2150.
https://www.nextbigfuture.com/2018/12/world-population-goes-up-at-most-500-by-2150-but-can-already-feed-those-people.html
Rather a lot of hand waving going on with generalities. Water shortages are already making some ag areas reduce productivity. There is no guarantee that some places, like Africa, can increase productivity due to political and legal issues. Climate change is going to put a lot of stress on crop yields. Factory meat is coming, but prices have to decline to make this substitute work.
People do not live on food alone. They need other resources – shelter, heat (or A/C), goods and services. We are already at overcapacity based on post-industrial society consumption. 40 bn people at these standards will break the planet very quickly.
next Big Future may be fun to read, but don’t take their articles too seriously.
The population won’t get that high.
When countries become prosperous people have fewer children and it’s clear that most people want to be as prosperous as First World, even Second World people.
The problem is gangster leaders who promise miracles while repressing people into virtual slavery,
Either millions will die from disease and poverty or there will be revolutions that will free countries to follow examples of prosperity. These may be leaders getting smart enough to alter their economies while maintaining power or leaders and their supporters being slaughtered like livestock at great expense to everyone.
But the population will either way not be that huge.
The complexity of such analyses is daunting but Mr. Trolley has made it understandable. I read somewhere that Super Earths may be more conducive to the development of life (more tectonic activity to recycle carbon, stronger magnetic fields to protect the atmosphere). However, such planets may also have more frequent major geological events resulting in major changes in the atmosphere throwing life for a loop as described in the article. Perhaps the activity is so often and intense that life gets knocked back to the single cell level on a regular basis. Earth may be lucky; one in thousand by being large enough to have adequate tectonic activity but not too large to result in excessive geologic activity.
You’ve just come up with yet another rare Earth factor; correct planet size.
Turns out there are other analyses that suggest Super Earths may not have tectonic activity due to formation of a thick rigid crust:
https://www.sciencenewsforstudents.org/article/super-earths-not-place-plate-tectonics
And this article suggests that tectonic activity may be crucial for long term evolution of complex life:
https://www.scientificamerican.com/article/earths-tectonic-activity-may-be-crucial-for-life-and-rare-in-our-galaxy/
Of course, the discussion is limited to Life As We Know It. I am partial to the Rare Earth hypothesis but our knowledge is just too limited to draw conclusions. Ask me in about a hundred years and I may have a stronger opinion:)
May yes, may be not, may be mini-Earth?
We do not have an answer, Mr. Tolley article is about our planet, not about something virtual “may be”, I sure our Universe is wider than our imgination, only space exploration can give the right answer, there is not second Earth, but may be there is lot of planets that are much “better” than Earth for the life forms development.
One important point is that after every large extinction event something better has come along. Evolution may need the chaos created by these large changes in nature to jump to a higher and more sophisticated level. I have not seen any long outcome in what was developed after the events that was a step or a major set back to evolution. This makes me wonder if life was brought here via panspermia
We should be aware of the fallacy of thinking that evolution keeps throwing up advances to some ultimate peak – us. What does appear to happen is that when some dominant groups are wiped out by change, another group emerges. We think of mammals as being more “advanced” than dinosaurs, but that is not necessarily true. It just happens that we have descended from the mammalian line. However, that is not a given, just the result of what we see with the 5 major extinctions teh fossil record records.
I’m thinking more in the line of chaos theory of complicated systems and when thrown out of whack jump to a higher level. Specifically DNA and its ability here or anywhere else to overcome obstacles. You might say it has a mind of its own!
Complex systems do not have to be chaotic. They can be more stable due to negative feedbacks. For chaotic systems, I am not aware that perturbations “jump to a higher level”, whatever that means in actuality. Do you have a reference for this statement?
“The universe is slightly tilted toward self-organizing processes, and these processes—as Prigogine was the first to elaborate—escape present-level turmoil by jumping to higher levels of self-organization.”
I disagree with this in that the universe is not slightly tilted toward self-organization but is self-organization in all of attributes. After a major extinction evolution will go into overdrive to fill and improve the ecological structure that was destroyed. Any surviving lifeform that was on the edge because of competition will then evolve from their higher level systems that developed to compete.
Good reads:
Turbulent Mirror by John Briggs & David Peat
Exploring Complexity by Gregoire Nicolis & Ilya Prigogine
The Earth is not ” Rare”.
It is unique.
But in terms of life as we know it , and indeed life as we don’t know it too, this means nothing in the greater meaning of things. Life could be ubiquitous and be seemingly perfectly adapted to whichever planet ( or moon et al ) it formed on . No doubt if that life is intelligent too it might well be describing its own home planet as rare too. ( I would like to think however that it would be better informed than the parochial crudely disguised retro Ptolemaic-cum-intelligent design vision that “Rare Earth” has come to symbolise to many of its chief exponents ). One thing is obvious from all papers such as this . Which is that a near innumerable number of factors have to interact at set points and over set times to form and maintain life. In finely balanced yet unstable equilibrium. Extend that over near immeasurable time and we are well into a position whereby simulations , and especially simulations based on a sample size of ONE, dear old “Rare Earth”, quite literally break down .
In chaos .
Agree, Unique – it is correct definition.
Indeed, we can use the simple game of plinko as a analog for the danger of drawing conclusions from a sample size of one. A single plinko piece could look back on its path and find events that are the product of extremely distinct conditions. If that plinko piece is unable to find other pieces and the plinko piece is so inclined, it could claim the entire distribution curve for itself and its path as the only path.
I take the point Ashley, AlexT and Harold that Earth is unique. But isn’t that semantics? Something that is unique would also be rare, no?
Life could be common or uncommon, we just don’t know yet. The way I see it though, there are only two possible pathways yeilding ubiquitous life; (1) Abiogenesis is easy, or (2) Life has been seeded widely. If you think life is common, take your pick. However, decades of lab results suggest (1) is very far from easy.
That may be like saying that baking a cake is not easy when experiments taking just seconds fails to create one. Life may need the right ingredients, container, “oven temperature”, and time to bake or simmer until done. It isn’t a magic trick. ;)
According to Wikipedia, in the Permian CO2 was 900 ppm but in the Ediacaran it was five times higher at 4500 ppm. So if the Siberian Traps had occurred in the Ediacaran, CO2 would have only marginally increased and temperatures would have risen only slightly.
The super continent of Panagea had already formed before the Siberian Traps so the land localized on one side of the planet affects the climate and the weathering on mountains which helps remove the CO2. The Appalachian mountains began at that time, but other mountain ranges are more youthful and formed after Panageas break up and collision of plates. There have been more than one super continient to due continental drift over earths history which affected the past climate. Earths polar ice caps began to form fifty million years ago in the Cenozoic, Paleiogene, Eocene.
The question is how long did it take for 4500 parts per million to be reduced to 900 parts per million? The Ediacaran period is is 635 to 541 Mya after the snowball Earth period. The volcanism and Co2 is what melted the snowball Earth and began the Cambrian explosion. Oxygen based life helped remove the Co2 with the carbon cycle and Urey reaction. 4500 parts would be catastrophic to our ecosystem today with only a five degree temperature. The Co2 was four times higher 100 million years ago and there were no polar ice caps.
I remember seeing an exhibit and the Academy of Sciences in San Francisco which was about the mass extinctions in the north polar region and arctic circle in Eocene fifty million years ago. There were subtropical temperatures, plant and animals there adapted to that environment and it’s warmer temperatures which became extinct with the drop in temperature . This occurred in only fifty million years and drop in Co2 levels.
Slightly off topic but with an enormous world ocean, very warm water and a faster rotation rate of the Earth in that era (intensified Coriolis effect), hurricanes must have been absolutely titanic in size and intensity with a high frequency of occurrence. As a fan of extreme meteorology, my imagination is in overdrive.
Google ‘Hypercane’ to see just how titanic they could have been!
Yes! The water temperature may be not have been 120 F back then but with plenty of room and a stronger Coriolis effect, the storms would have been stupendous.
My point was the super continent Pangea had only one mountain range or they were only on one side of the planet which with all the land on one side of the planet. Also the temperatures on all places over the entire globe were warm. The south pole only one hundred million years ago was an estimated 65 degrees and it would be even hotter there during the Ediacaran. It does not take a large temperature increases harm the ecosystem.
The cause of most if not all of the mass extinction are caused by major impacts in the history of the earth.
Antipodal hotspots and bipolar catastrophes: Were oceanic
large-body impacts the cause?
Abstract
“One aspect of the hotspot distribution that has received little attention is its antipodal character. Of 45 dprimaryT hotspots
found in most hotspot compilations 22 (49%) form antipodal pairs within observed hotspot drift limits (V 20 mm/yr). In
addition, the available ages, or possible age ranges, for both hotspots of an antipodal pair tend to be similar (V 10 Myr difference) or overlap. Monte Carlo simulations indicate that the antipodal primary hotspots’ locations and ages are not due to chance at the N 99% confidence level ( p b0.01). All hotspot pairs include at least one oceanic hotspot, and these are consistently opposite those hotspots related to large igneous provinces (LIPs) and continental volcanism. A mechanism of formation is considered in which minor hotspot volcanism is induced at, and flood basalt volcanism is triggered by seismic energy focused antipodal to, oceanic large-body impact sites. Because continental impacts are expected to have lower seismic efficiencies, continents possibly acted as shields to the formation of antipodal hotspot pairs. Published numerical models indicate that large oceanic impacts (10-km-diameter bolide) generate megatsunami capable of altering coastal depositional
environments on a global scale. Past impact-generated megatsunami, consequently, could have left widespread stratigraphic records, possibly misinterpreted as indicating large rapid changes in eustatic sea level, and widely disrupted continental and marine sediment reservoirs responsible for abrupt changes in the isotopic composition of seawater. Phanerozoic mass extinction events, therefore, might have resulted primarily from catastrophic megatsunami in a dominantly oceanic hemisphere and the near contemporaneous effusion of vast quantities of noxious gases from flood basalt eruptions in a dominantly continental one.”
http://www.mantleplumes.org/WebDocuments/Antip_hot.pdf
LARGE SCALE IMPACTS AND TRIGGERED VOLCANISM.
“The idea of impact induced volcanism
continues to blossom. However, this appealing idea is seldom supported with an appropriate physical mechanism. The aim of this
publication is to critically examine some frequently cited mechanisms of impact energy transformation into a trigger for terrestrial volcanism and magmatism.”
https://www.lpi.usra.edu/meetings/largeimpacts2003/pdf/4062.pdf
GRACE gravity evidence for an impact basin in Wilkes Land, Antarctica.
Abstract
“New details on the east Antarctic gravity field from the Gravity Recovery and Climate Experiment (GRACE) mission reveal a prominent positive free?air gravity anomaly over a roughly 500?km diameter subglacial basin centered on (70°S, 120°E) in north central Wilkes Land. This regional inverse correlation between topography and gravity is quantitatively consistent with thinned crust from a giant meteorite impact underlain by an isostatically disturbed mantle plug. The inferred impact crater is nearly three times the size of the Chicxulub crater and presumably formed before the Cretaceous formation of the east Antarctic coast that cuts the projected ring faults. It extensively thinned and disrupted the Wilkes Land crust where the Kerguelen hot spot and Gondwana rifting developed but left the adjacent Australian block relatively undisturbed. The micrometeorite and fossil evidence suggests that the impact may have occurred at the beginning of the greatest extinction of life on Earth at ?260 Ma when the Siberian Traps were effectively antipodal to it. Antipodal volcanism is common to large impact craters of the Moon and Mars and may also account for the antipodal relationships of essentially half of the Earth’s large igneous provinces and hot spots. Thus, the impact may have triggered the “Great Dying” at the end of the Permian and contributed to the development of the hot spot that produced the Siberian Traps and now may underlie Iceland. The glacial ice up to a few kilometers thick that has covered the crater for the past 30–40 Ma poses formidable difficulties to sampling the subglacial geology. Thus, the most expedient and viable test of the prospective crater is to survey it for relevant airborne gravity and magnetic anomalies.”
https://agupubs.onlinelibrary.wiley.com/doi/full/10.1029/2008GC002149
A Possible Cause of the Siberian LIPs: “Collisions
Aggrgation Effect” of an Aerolite Impact .
Abstract: “The authors believe that the main reason for the formation of the Siberian LIPs may be the nonlinear “Collisions
Aggregation Effect” caused by a meteorite impact in the Falkland Plateau crater or Wilkes Land crater, and the correspondence of
the Falkland Plateau crater may be better. As a result, the authors have reached the following conclusions and understandings: 1.
The late Permian: the siberian LIPs are in the Siberian plate at 60°N, 80°E; the Falkland Plateau crater is located in the
southwestern corner of the Gondwana, about at 70°S, 30°E, at the southernmost point of the South American plates, close to the
southern orogenic belt of the South America and the stress state of the Falkland Plateau crater may be under pressure; the Wilkes
Land crater may be located at 60°S, 110°E, at the eastern Antarctic continent, close to the Australian plate, is not located in the
orogenic belt and the stress state of the Wilkes Land crater should not be pressure-type, but is in a wide range of ice covered area;
2. Figure 6 and 7 show the location relationship between the Siberian LIPs and Falkland Plateau crater or Wilkes Land crater does
not have a antipodal characteristic, nor have a linear “Collisions Aggregation Effect” profile, but have a nonlinear “Collisions
Aggregation Effect” distribution, the Falkland Plateau crater correspondence may be better; 3. Whether or not the Falkland
Plateau crater or Wilkes Land crater was finally confirmed as a true meteorite crater, their relationships with the Siberian LIPs
should have a nonlinear distribution of “Collisions Aggregation Effect” when the time they formed close to the Siberian LIPs,
and is the main reason for the formation of the Siberian LIPs. The velocity vector direction of the meteorite impact may have a
smaller angle of impact, and significantly eastward (relative to the Earth’s center); 4. About the celestial body which its impact
can form LIPs or cause the formation of LIPs, it should have: the impact time should be earlier than LIPs or almost
simultaneously; a certain size; the “impact point” and the LIPs distribution area have the “Collisions Aggregation Effect”
distribution characteristics; 5. Figure 2 shows a possible formation model about LIPs of a celestial body collision: due to the
violent impact of a celestial body, created a tectonic weakness in the crust, even up to the crust-mantle boundary, a fault zone
pointing to the Earth’s center, and a radial spherical fracture group at the “impact point” or “collisions aggregation point”; the
magma originating from the mantle intrude along the weak zone under pressure, a large number of magma invaded and gathered
in the radial spherical fracture group, causing the overlying strata rising slowly, forming a dome and a similar head-tail structure
of the mantle plume that the head dilated, tail slender deep into the mantle on the longitudinal.”
http://article.sciencepublishinggroup.com/pdf/10.11648.j.earth.20180703.12.pdf
Whatever the initial event that resulted in teh Permian extinction, the data indicates that warming was the decisive cause, rather than any event that may have triggered the volcanism.
However, this raises an interesting question. If we had major extinctions, whose initial cause was a common cosmic event – i.e. asteroid impacts – then we must expect that this is a common feature in other star systems. IOW, we should expect most rocky exoplanets to bear scars of asteroid impacts and that any living worlds would have been repeatedly impacted by such asteroids, disruption their biospheres.
One reason the Earth may have been affected by comets and asteroids impacts more often than in other systems is that Jupiter acts as a collector of comet and asteroid families. One of my favorite books is Monsters in the Sky, by Paolo Maffei and I recommend everyone should read it. Even though written in 1976 this book is still current and does very well at explaining and illustrating such things as Jupiters comet families and their evolutions.
As to other systems, M Dwarfs and planet in resonant orbits around them, the low metalicy and non Jupiter size planets would tend to support more stable and possible more organic features on these worlds.
“One reason the Earth may have been affected by comets and asteroids impacts more often than in other systems is that Jupiter acts as a collector of comet and asteroid families”.
Interesting, because in the past (and still?) Jupiter was often considered a great protector of the Earth by attracting many potential impactors.
Others have indeed argued that Jupiter probably slings about as many (if not more) impactors toward Earth as it sucks up.
Which is more the case: Jupiter as a impact protector, or Jupiter as an impact enhancer?
I suppose those two opposite argument are convenient to use to prove any point of view.
It is far from science, but mostly working like “dice throws”, bet what you want every case has the same probability.
As usual the Devil’s in the details:
Jupiter: Friend Or Foe?
“The traditional belief that Jupiter acts as a celestial shield, deflecting asteroids and comets away from the inner solar system, has been challenged by the first in a series of studies evaluating the impact risk to the Earth posed by different groups of object. Astronomers now note the impact hazard posed to Earth by the Centaurs, the parent population of the Jupiter Family of comets. The presence of a Jupiter-like planet in the solar system does not necessarily lead to a lower impact rate at the Earth.”
The problem is that they talk of a smaller planet then Jupiter causing more earth impacts but do not go on to include Saturn, Uranus and Neptune in the calculations.
https://www.sciencedaily.com/releases/2007/08/070824133636.htm
And another for Foe;
Jupiter:
friend or foe? An answer.
Barrie Jones and Jonti Horner summarize the results of models addressing the role of Jupiter in protecting – or otherwise – the Earth.
“Conclusions The idea that the planet Jupiter has acted as an impact shield through the Earth’s history is one that is entrenched in planetary science, even though little work had been done to examine this idea. In this work, we detail the results of simulations that reveal that Jupiter’s in?uence is not so straightforward. Indeed, it seems that the presence of Jupiter actually increases the rate at which asteroids and short-period comets impact the Earth. The traditional idea of “Jupiter – the shield” only holds true when one considers the long-period comets, which are so ef?ciently ejected from the solar system as Jupiter gains in mass that few remain to threaten the Earth. Given that these comets only make up a small fraction of the total impact threat, our startling conclusion is that, overall, Jupiter is not friend but foe! ”
http://www.jontihorner.com/papers/FoFAnswer.pdf
Jupiter’s Protective Role Questioned.
https://centauri-dreams.org/2012/02/20/jupiters-protective-role-questioned/
On balance , Jupiter – friend. Along with Saturn too.
Not by acting as Zeus’s shield Aegis though. First by acting as a hoover to mop up the nascent Sun’s protoplanetary disk within just a few million years . Then later blowing away much of the remainder in going walkees big time early on in Solar system history . It’s “Grand Tack” cum-giant-jumping-Jupiters routine choreographed in a resonant push me pull me dance with Saturn acted like the mother of all brooms . Clearing out vast amounts of leftover detritus that might otherwise still be around to pummel the Earth today. This is no more evident than in the very low reactive masses of both the asteroid and Kuiper belts . A further marker of this is in the background solar system dust level , the erstwhile “zodiacal light” , which is also far lower than for most if not all other stars observed to date .
This may enlighten you to why and reasoning of a Comet called Comet Encke or Encke’s Comet (official designation: 2P/Encke) may have been the creator of the end of the ice age world in 12,900 BP. This could be happening again in the presence of multiple meteor streams within the Taurid Complex, including “asteroids” orbiting within them, indicates that the original comet evolved via a cascading hierarchy of fragmentations.
Astronomical Hypothesis for the YDB Impact Event.
“The fragments of a splitting event disperse as they move away from the comet. Recently released cometary material forms an elongated, dense trail typically a few hundred Earth radii long and 10 radii wide within an orbit (Napier 2015; Napier et al. 2015). Taking account of orbital precession and nutation, the recurrence time of encounters between Earth and one such debris swarm in an Encke-like orbit is t ? 500/j My, where the effective cross-sectional area j of the swarm is in Earth radii. If there is an average of one such swarm at any time, we expect passage through it about once in 50,000 y, in the course of which the Earth will encounter 1017/j ? 1013–1014 g of material over a few hours, entering the Earth’s atmosphere at 30 km/s. The debris so encountered will generally be a mixture of dust and larger frag- ments, energetically equivalent to the impact of
?1000–10,000 Tunguskas and with the potential to create severe biotic and climatic disturbances (Hoyle and Wickramasinghe 1978).”
?1000–10,000 Tunguskas type airburst or impacts is something we are not ANYWHERE near term capable of dealing with!!!
Centaurs as a Hazard to Civilization (2015)
https://www.buckingham.ac.uk/wp-content/uploads/2011/09/Centaurs-as-a-hazard-to-civilisation.pdf
Are asteroids hiding among the Taurids?
https://earthsky.org/astronomy-essentials/taurid-new-meteroid-stream-potentially-hazardous-asteroids
So if you wish upon a bright Taurid fireball in mid October, just remember it might be civilizations doom…
https://cosmictusk.com/astronomical-hypothesis-younger-dryas-boundary-impact-event/
Incoming! A June meteor swarm could be loaded with surprises.
“Scientists studying a mysterious event over Siberia in 1908 call for a special observation campaign next summer.”
“On June 30, 1908, an object the size of an apartment building came hurtling out of the sky and exploded in the atmosphere above Siberia. The Tunguska event, named for a river, flattened trees for 800 square miles. It occurred in one of the least-populated places in Asia, and no one was killed or injured. But the Tunguska airburst stands as the most powerful impact event in recorded human history, and it remains enigmatic, as scientists don’t know the origin of the object or whether it was an asteroid or a comet.
One hypothesis: It was a Beta Taurid.
The Taurids are meteor showers that occur twice a year, in late June and late October or early November. The June meteors are the Betas. They strike during the day, when sunlight washes out the “shooting stars” that are visible during the nighttime meteor shower later in the year.
A new calculation by Mark Boslough, a physicist at Los Alamos National Laboratory, shows that the tree-fall pattern in Siberia is consistent with an asteroid coming from the same area in the sky as the Taurid meteor swarm. Boslough and physicist Peter Brown of Western University in London, Ontario, gave a presentation at the American Geophysical Union fall meeting in Washington this month in which they called for a special observation campaign this June to search for Tunguska-class or larger objects embedded in the Taurids.
In some years, Earth passes near the densest cluster of material in the Taurid stream — and 2019 will be such a year. The scientists say it presents potentially the richest batch of incoming material since 1975, when seismometers left on the moon by Apollo astronauts recorded a spike in impacts during the Taurid swarm.”
“This ring of material is roughly but not exactly on the same plane as the Earth’s orbit. That means the Earth crosses the Taurid stream twice a year. The June crossing intersects Taurid material traveling away from the sun, and the October crossing intersects material traveling toward the sun. That’s why you can see the October Taurids as they hit the Earth’s atmosphere. The June Taurids are washed out by sunshine but can be spotted by radar.”
The June Taurids are part of the Comet Encke fragmentation and possible what is left of a much larger original super size Comet. It would be interesting to see if this is something that the amateur astronomer community could participate in with astrophotography…
https://www.washingtonpost.com/amphtml/science/2018/12/25/incoming-june-meteor-swarm-could-be-loaded-with-surprises/
Taurids can be a source of cosmic disaster.
27.01.2017
Observations of Polish astronomers working in the Polish Fireball Network (PFN) show that in the Taurids there is a narrow stream of cosmic bodies, the largest of which, in the event of a collision with Earth, can cause a disaster on a continental scale. “Moreover, this stream does not tend to disperse over time, but it is maintained stably by the resonance with the orbit of Jupiter. That is not the end of bad news. In 2019 there will be another Earth\’s meeting with the resonance stream. This time it will be a daytime encounter, at the turn of June and July – just like during the Tunguska event. As if that were not enough, the encounter will be even closer than during the very high and bolide-intensive years 2005 and 2015 “- told PAP Prof. Arkadiusz Olech from the Nicolaus Copernicus Astronomical Centre PAS in Warsaw.
Scientists from the Polish Fireball Network have identified as many as 10 asteroids with sizes ranging from tens to hundreds of meters, which have almost the same orbits as Taurids. “And if Taurids enter our atmosphere, causing the phenomenon of meteors, nothing stands in the way of an encounter with a larger object. Especially that the Tunguska event, which was caused by an object with the size of about 50-100 meters, clearly shows that such things can happen” – explained Dr. Olech.
http://scienceinpoland.pap.pl/en/news/news%2C412823%2Ctaurids-can-be-a-source-of-cosmic-disaster.html
Discovery of a new branch of the Taurid meteoroid stream as a real source of potentially hazardous bodies.
https://www.aanda.org/articles/aa/pdf/2017/09/aa30787-17.pdf
Palaeolithic extinctions and the Taurid Complex.
Monthly Notices of the Royal Astronomical Society.
2 A LARGE PALAEOLITHIC SHORT-PERIOD COMET
2.1 The zodiacal cloud and the Taurid meteors
2.2 Near-Earth objects associated with Comet Encke
3 EVOLUTION OF A LARGE DISINTEGRATING COMET.
4 SOME GEOPHYSICAL EXPECTATIONS.
4.1 Thermal effects
4.2 Nanodiamonds
4.3 Lunar debris
https://academic.oup.com/mnras/article/405/3/1901/966774
https://academic.oup.com/mnras/article-pdf/405/3/1901/2894095/mnras0405-1901.pdf
I wonder if this Taurid object stream could be the cause of that meteor crater recently found in Greenland?
Well, that’s what the general consensus seems to be pointing at:
The Cosmic Tusk has a lot of info. https://cosmictusk.com/
And what is surprising is that every time I look, there is new and interesting material;
https://cosmictusk.com/alaska-muck-hibben-frozen-mammoth-comet-catastrophe/
Bill Napier seems to be the one that came up with the original idea that the Beta Taurid stream being the producer of these impacts. But there apparently has been a long history going back at least 80 years relating to the Alaska and Yukon muck and what created it.
Some of the prehistoric and even historical tsunami evidence around the pacific rim could be cause from impacts instead of earthquakes.
But there is plenty of material out there on the internet on the subject.
More craters under Greenland’s glaciers!!!
ttps://cosmictusk.com/another-crater-hiawatha-greenland-joe-macgregor_steve_garcia/
https://cosmictusk.com/another-crater-hiawatha-greenland-joe-macgregor_steve_garcia/
Kinda off topic, but we should also consider an artificial silence of the bugs scenario too; without a single doubt it will affect the food chain. Lots of people think we will use bugs as a source of meat in the future but this case seems longer feasible to feed 9-10 billions people, maybe lab grown meat?
Lab grown meat is almost certainly in our future if the price becomes competitive. There is no question that beef eating is a significant contributor to AGW, and all meats from farmed animals and poultry have a higher carbon footprint than plant crops. I could see that humans might become disgusted by the thought of eating the flesh of animals as well, making animal farming a declining activity. (Few people will eat raw meat, and many folks have trouble eating unusual animals or protein sources. So we are moving in that direction.) I’ve eaten a tofu chicken substitute that was quite good, although the price was ridiculously high and suited to the affluent only. We’ll see where lab-grown meat goes in the next decade. As we have seen with fossil fuel interests blocking renewables, the US cattlemen associations are already lobbying to make lab meats difficult to produce and sell. Let’s hope we don’t look forward to Soylent Green day.
I’m thinking like the difficulty of having fresh meat in space especially during a long (human) space mission ( to Mars, Jupiter’s moon-Europa etc…) and raising cows or pigs in space is a big no no, hence the possibility of bringing a certain type of insect eggs as a food source or lab grown meat, not sure which one is more disgusting.
This video from 2012 says that NASA still has yet to achieve feeding a human crew on a manned Mars expedition for the several years it would take to travel to and from the Red Planet, without a resupply along the voyage. Is this still the case?
https://www.youtube.com/watch?v=GGmaXX59aq8&feature=youtu.be
The issue is “without a resupply”. Why on earth (or Mars?) would anyone assume that resupply was not an option? Zubrin’s Mars Direct approach allows for propellant resupply by in situ manufacture. I seriously doubt that we cannot keep food dried or frozen for years – we already do that on Earth. So an early food dump on Mars to ensure that adequate food is available is an option – it’s basic logistics.
They were referring to early expeditions that would visit Mars and return before starting colonization. However your point is valid.
Going to Mars will not be like going to the Moon with Apollo and not just because of the extra distance and time involved.
About the disgusting part, the novella The State Of The Art mentions briefly of using human cells to grow “your own meat” during the grand party before the good old ship Arbitrary leaving our Solar System. This is so gross that it should be illegal using human cells to produce “special lab grown meat”.
Anyway, if a similar event happens, it will likely take less than 1000 years all thanks to the artificial acceleration. This is still too early to know about exactly what is going on but most likely some group of people want to scare the public in order to do something to the global climate change.
Re: “Rare Earth”.
I agree with physicist Paul Davies (The Eerie Silence) that we just do not yet know whether we are an anomaly or the universe is full of life. I think we will have strong hints from spectroscopic atmosphere data of exoplanets in a decade or so. These proxy data will not be definitive, but suggestive, either way. As to the question of whether life arises from abiogenesis wherever it is found, or whether it is the result of panspermia, we will not know without samples. That we can only determine from our own solar system, although interstellar wanderers like ‘Ouuamua many also provide possible answers.
If Davies’ hypothesis of a “shadow biosphere” on Earth was ever to be confirmed, then that would lead to a strong conclusion that abiogenesis is common.
Antipodal volcanism at the Siberian Traps is not the only material brought up from an impact on the opposite side of the earth. The ‘Deep Biosphere’ in Deep Earth Totals 15 to 23 Billion Tonnes of Carbon most of it in oil, gas and coal and may go much deeper than anybody thought. Burning oil, gas and coal may have been what caused the demise of so much surface life 252 mya.
Map, courtesy of Jo Weber, showing the extent of the Siberian Traps as based upon estimates derived from Masaitis (1983). Circles show major gas fields; diamonds show major oil fields; data from Horn (1983)
https://www.researchgate.net/profile/J_Herndon/publication/305630194/figure/fig7/AS:391356854882321@1470318153523/Map-courtesy-of-Jo-Weber-showing-the-extent-of-the-Siberian-Traps-as-based-upon.png
Could Life on Mars Be Lurking Deep Underground?
https://www.livescience.com/64318-mars-life-deep-biosphere.html
Scientists Reveal a Massive Biosphere of Life Hidden Under Earth’s Surface.
https://www.sciencealert.com/scientists-lift-lid-on-massive-biosphere-of-life-hidden-under-earth-s-surface
Life in Deep Earth Totals 15 to 23 Billion Tonnes of Carbon—Hundreds of Times More than Humans.
https://deepcarbon.net/life-deep-earth-totals-15-23-billion-tonnes-carbon
The Deep Hot Biosphere: The Myth of Fossil Fuels.
“always original, always important, usually controversial, and usually right”
– FROM THE FOREWORD BY FREEMAN DYSON
“When scientists discovered thermophiles?primitive microorganisms that live in deep seafloor vents and eat hydrocarbons (chemicals like gasoline)?experts assumed the mysterious bugs had little to tell us about ourselves or about the earth’s core. Cornell University Professor Emeritus Gold, however, who for 20 years directed the Cornell Center for Radiophysics and Space Research, here proposes the striking theory that “a full functioning… biosphere, feeding on hydrocarbons, exists deep within the earth, and that a primordial source of hydrocarbons lies even deeper.” Most scientists think the oil we drill for comes from decomposed prehistoric plants. Gold believes it has been there since the earth’s formation, that it supports its own ecosystem far underground and that life there preceded life on the earth’s surface. The “deep hot biosphere” hypothesis would explain the thermophiles, the minerals and the oil Swedish drillers found in 1990 under rock where no one expected them. The hot goo and massed gas far under our feet would also explain some mysterious historical earthquakes (notably the New Madrid, Mo., shocker of 1811), and it would tell puzzled geologists why so many oil reserves just happen to sit underneath coal fields. As later chapters explain, if Gold is right, the planet’s oil reserves are far larger than policymakers expect, and earthquake-prediction procedures require a shakeup; moreover, astronomers hoping for extraterrestrial contacts might want to stop seeking life on other planets and inquire about life in them.”
Seawater Can Reach Right To The Edge Of The Earth’s Core.
“Underwater lava flows reveal that seawater gets cycled to depths within the Earth as great as 2,900 kilometers (1,800 miles), the boundary between the mantle and the outer core. The discovery of seawater trapped in basalt not only overturns theories about what happens when tectonic plates are forced beneath each other, but provides evidence the oceans and atmosphere that make life possible come from comets or icy meteorites.”
https://www.iflscience.com/physics/seawater-can-reach-right-to-the-edge-of-the-earths-core/all/
There’s as much water in Earth’s mantle as in all the oceans.
https://www.newscientist.com/article/2133963-theres-as-much-water-in-earths-mantle-as-in-all-the-oceans/
So were there is water, usually there is life, deep very deep in the Earth!
And Mars and Bennu, Yes Bennu the perfect place for life to hide in a wet asteroid, where no cosmic rays or solar storms can reach them…
There are limits. Organic molecules fall apart with heat. No organism survives at more than 122C (so far), and I cannot see how carbon will hold together at extreme temperatures. With the Earth temperature profile of 15-30C/km increase, this means that the maximum depth is about 8km. 2 boreholes at 9km did not find life, and the deepest borehole at 12km found water and microfossils, but not living organisms. Certainly, there is no life in the mantle, at least no life as we think we know it.
I have proof! All those beautiful crystal Geodes are actually fried Dragons eggs from inner earth! ;-)
Tremendously interesting article and comments. Brilliantly done Alex.
Patient Observer, I am a fan of extreme meteorology too. My interest in meteorology got me into planetary science early. It is true that a fast rotation and strong Coriolis effect will create larger and more powerful storms. There were much more powerful hurricanes during the Permian extinction because of the higher temperatures or 10 degree temperature increase. A higher temperature results in more evaporation of sea water and higher percentage of water vapor in the atmosphere and more rainfall. Warm air holds more water vapor than cold air as proven by the condensation law, relative humidity and the adiabatic lapse rate 5.5 degrees per 1,000 feet of rise, so with higher temperature there are stronger hurricanes.
Over a long period of time the rain will remove the CO2 from the air through the Urey reaction and carbon cycle. The carbonic acid in the ocean would eventually be removed.
An Earth twin exoplanet which was a little closer to the Star than Earth might be pushed to a temperature too high for life which would become extinct or nearly extinct. It might be harder for life to start over.
What could have wiped 3km of rock off the entire Earth?
“Believe it or not, the geology at the bottom of the Grand Canyon is extraordinarily common. There, layers of sedimentary rock lie flat atop angled layers of significantly more ancient metamorphic rock. The gap there is enormous—if Earth’s rocks constitute a book of the planet’s history, there are about a billion pages missing. The story only picks up again around 540 million years ago in the Cambrian period, with an evolutionary explosion of complex life just as remarkable as the sudden change in the rock.
This gap can be found all around the world, and has picked up the name the Great Unconformity. Cambrian sedimentary rocks rarely rest on anything other than much older metamorphic or igneous rock, implying that whatever rock formed in the intervening time was scrubbed away by something. This erasure of a chunk of geologic history has long been an enticing mystery for geologists.”
https://arstechnica.com/science/2019/01/huge-break-in-geological-record-could-be-due-to-a-snowball-earth/
Earth is missing a huge part of its crust. Now we may know why.
A fifth of Earth’s geologic history might have vanished because planet-wide glaciers buried the evidence.
For one thing, “around 600 to 700 million years ago, Earth loses its craters,” notes study coauthor Bill Bottke, a planetary scientist and asteroid expert at the Southwest Research Institute in Boulder, Colorado. Some ancient craters still exist on stable continental cores named cratons, but they are few and far between.
https://www.nationalgeographic.com/science/2018/12/part-earths-crust-went-missing-glaciers-may-be-why-geology/
Seems we still have a lot to learn…
Did life on land arrive far earlier than thought?
A new analysis of fossils from Earth’s oldest soil suggests that multicellular, land-dwelling organisms possibly emerged much earlier than thought.
The study, published in the January 2019 issue of the journal Sedimentary Geology, looked at fossils previously considered to be ocean organisms. These fossils were found in sediments in South Australia that date to between 542 million to 635 million years ago – during a geological period known as the Ediacaran.
Greg Retallack, fossil collections director at the University of Oregon’s Museum of Natural and Cultural History, is the study lead author. He said in a statement:
These Ediacaran organisms are one of the enduring mysteries of the fossil record. Were they worms, sea jellies, sea pens, amoebae, algae? They are notoriously difficult to classify, but conventional wisdom has long held that they were marine organisms.
But new research – based on a geochemical and microscopic re-examination of the thin, silty-to-sandy layers of sediments where the fossils were found – suggests otherwise.
https://earthsky.org/earth/life-on-land-arrive-far-earlier-than-thought