Centauri Dreams
Imagining and Planning Interstellar Exploration
A Landing Site for Dragonfly
Rotorcraft have certainly been in the news lately, with Ingenuity, the Mars helicopter, commanding our attention. The Dragonfly mission to Titan involves a far more complex rotorcraft capable of visiting numerous destinations on the surface. In fact, Dragonfly makes use of eight rotors and depends upon an atmosphere more helpful than what Ingenuity has to work with on Mars. Titan’s atmosphere is four times denser than what we have on Earth, allowing Dragonfly to move its entire science payload from one location to another as it examines surface landing zones while operating on a world whose gravity is but one-seventh that of Earth.
I want to call your attention to the publication of the science team that just appeared in the Planetary Science Journal, because it lays out the rationale for the various decisions made thus far about operations on and above Titan’s surface. It’s a straightforward, interesting read, and makes clear how much work we have to do here. Yes, we had Cassini for a breathtaking tour that lasted 13 years, with repeated flybys and investigations on Titan using radar, but while we know a lot about structures like lakes and mountains on the surface, we know all too little about their composition.
In fact, as Alex Hayes points out, we didn’t know at the time Cassini launched whether the Huygens probe would find a global ocean at Titan or a solid surface of ice and organics. Because of the uncertainty, the Huygens science experiments were primarily atmospheric, meant to function during the descent phase. Hayes (Cornell University) is a co-investigator for Dragonfly. He adds:
“The science questions we have for Titan are very broad because we don’t know much about what is actually going on at the surface yet. For every question we answered during the Cassini mission’s exploration of Titan from Saturn orbit, we gained 10 new ones.”

Image: What we do know. This is Figure 1 from the paper. Caption: Dragonfly will image from the surface to provide context for sampling and measurements, as well as in flight to identify sites of interest at a variety of locations. (Left) Huygens image of Titan’s surface; cobbles are 10-15 cm across and may be water ice (Tomasko et al. 2005; Keller et al. 2008; Karkoschka & Schröder 2016a). (Right) Huygens aerial view of terrain akin to the diverse equatorial landscapes that Dragonfly will traverse and image at higher resolution. Credit: NASA/ESA/Barnes et al.
Dragonfly’s 2.7 year mission, starting upon arrival at Titan in 2034 during winter in the northern hemisphere, will commence at a landing site that was chosen for its safety factors (broad, relatively flat terrain) as well as its proximity to nearby interesting scientific targets. The goal is to set down at the equatorial dune fields called Shangri-La, which NASA notes are similar to the dunes found in Namibia on Earth. A series of short flights will explore this area before longer flights of up to eight kilometers begin, the beauty of the design being that Dragonfly will be able to sample interesting surface areas along the route to its destination, the Selk impact crater.
As the mission now stands, the lander should log on the order of 175 kilometers across Titan enroute to Selk. The latter is an interesting place because there is evidence here of past liquid water as well as organics, complex molecules containing carbon, along with hydrogen, oxygen and nitrogen. Methane rain and a snow of organics keep Titan’s weather systems complex amidst a landscape containing the building blocks of life.
But let’s get back to that landing site. The paper refers to Shangri-La as an “organic sand sea,” with the touchdown site located 134 kilometers south of Selk Crater, and approximately 175 kilometers north-northwest of the Huygens Landing Site. The image below is Figure 7 from the paper, giving the landing site in context.
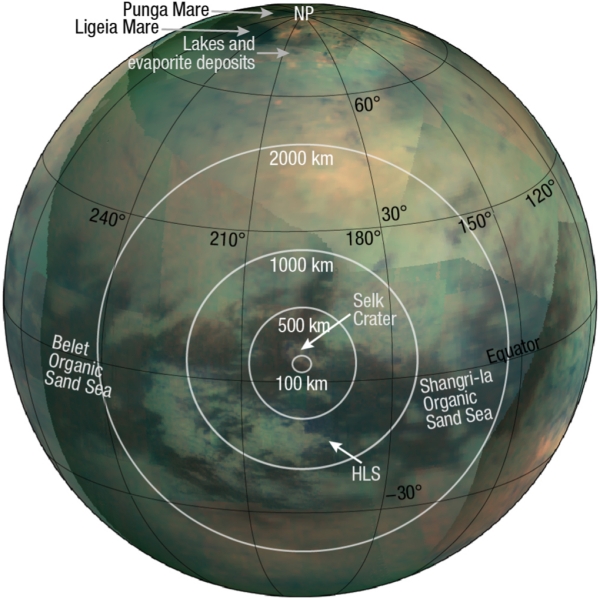
Image: Dragonfly landing site. Credit: Barnes et al.
As the paper notes, a ‘sand sea’ is only partially sand. Dunes can be separated by flat sand-free areas called ‘interdunes,’ a feature likewise common to Namibia, where the Namib sand sea is covered only 40 percent by sand. The interdunes that make up the balance are primarily gravel. Cassini was able to resolve Titan’s interdunes to reveal their predominantly icy character, one that matches the spectral properties at the Huygens landing site. The authors find the correlation interesting because it implies the Shangri-La interdunes will include water-ice gravels, “potentially a fine-grained layer damp with condensed methane,” and thus offering a chance for Dragonfly to sample both Titan’s organic sands and materials with a water ice component.

Image: This illustration shows NASA’s Dragonfly rotorcraft-lander approaching a site on Saturn’s exotic moon, Titan. Taking advantage of Titan’s dense atmosphere and low gravity, Dragonfly will explore dozens of locations across the icy world, sampling and measuring the compositions of Titan’s organic surface materials to characterize the habitability of Titan’s environment and investigate the progression of prebiotic chemistry. Credit: NASA/JHU-APL
The path to Selk Crater should take in a variety of terrain with different compositions, which will include the edge of the crater’s ejecta deposits. From Cassini data, the authors believe this material is similar in composition to the Huygens landing site, representing an area likely to feature water ice. Selk itself is 80 kilometers in diameter. Cassini data along with the Dragonfly team’s modeling show the spectral signature of organic sand in the interior and water-ice around the edges of the crater floor.
As you can see, the astrobiological examinations Dragonfly will engage in are both water-based and hydrocarbon-based, meaning a potential biosignature is possible from impact melt deposits or interactions with the interior ocean — this would be life as we know it — or from a form of life we have yet to discover that draws on liquid hydrocarbons within Titan’s lakes, seas and aquifers. The mission is designed around the ability to seek out both, as the paper explains:
We designed the science of Dragonfly around the themes of prebiotic chemistry, habitability, and the search for biosignatures, with an explicit consideration of both water and hydrocarbon solvents. To address prebiotic chemistry, we will determine the inventory of prebiotically relevant organic and inorganic molecules and reactions on Titan. In the realm of habitability, we will determine the role of Titan’s tropical atmosphere and shallow subsurface reservoirs in the global methane cycle, determine the rates of processes modifying Titan’s surface and rates of material transport, and constrain what physical processes mix surface organics with subsurface ocean and/or melted liquid-water reservoirs. Our search for biosignatures will entail a broad-based search for signatures indicative of past or extant biological processes.
The paper is Barnes et al., “Science Goals and Objectives for the Dragonfly Titan Rotorcraft Relocatable Lander,” Planetary Science Journal Vol. 2, No. 4 (full text).

Trajectory Watch: Charting Asteroid Bennu’s Future Path
In addition to its sample return mission at asteroid Bennu, OSIRIS-REx (Origins, Spectral Interpretation, Resource Identification, Security-Regolith Explorer) has tightened our projections about the object’s future trajectory. Although the impact possibility on Earth through the year 2300 is on the order of 1 in 1750 (0.057%), it’s an object we want to keep an eye on, because in 2135 Bennu will make a close approach to Earth that could nudge its trajectory in ways that are difficult to anticipate.
OSIRIS-REx spent more than two years working near the 500-meter wide asteroid, studying its mass and composition while tracking its spin and orbital trajectory. In terms of the latter, even factors as tiny as the force the spacecraft exerted during its sample collection event in October of 2020, a mere touch-and-go, had to be considered (the study confirms that the effect was negligible). Far more significant is the Yarkovsky effect, which occurs as solar heating eases on the nightside during the asteroid’s rotation and is radiated away as infrared energy, generating thrust.
It’s the tiniest of effects at Bennu, says Steve Chesley (JPL), a co-investigator on the study that has just appeared in Icarus, but of course it builds over time and has consequences for the asteroid’s path:
“The Yarkovsky effect will act on all asteroids of all sizes, and while it has been measured for a small fraction of the asteroid population from afar, OSIRIS-REx gave us the first opportunity to measure it in detail as Bennu traveled around the Sun. The effect on Bennu is equivalent to the weight of three grapes constantly acting on the asteroid – tiny, yes, but significant when determining Bennu’s future impact chances over the decades and centuries to come.”
Imagine how tricky it is to measure the cumulative effects of the Yarkovsky effect on a rotating object of uneven shape. Three close encounters in 1999, 2005 and 2011 were extensively tracked from the ground, but OSIRIS-REx data from the object itself have now allowed researchers to model an asteroid’s trajectory to the highest level of precision ever. Bennu’s future path is well known up to 2135. The question during the close encounter in that year will be whether it will pass through a dangerous ‘gravitational keyhole,’ an area where, as it responds to the effects of Earth’s gravity, the asteroid could become more likely to present an impact threat in the future. There is one ‘keyhole’ solution that would result in an Earth impact in 2182, with an impact probability of 1 in 2,700 (or about 0.037%).
In addition to the Yarkovsky factor, scientists have to consider the gravitational influence of the Sun and other objects in the Solar System including more than 300 asteroids that could have some effect. The solar wind streaming at variable rates outward from the Sun has to be considered as a source of pressure, and remember that OSIRIS-REx also discovered that Bennu was shedding rock particles, likely the result of thermal fracturing due to heating and cooling during rotation. These events along with the drag caused by interplanetary dust all affect the future trajectory.

Image: This mosaic of Bennu was created using observations made by NASA’s OSIRIS-REx spacecraft that was in close proximity to the asteroid for over two years. Credit: NASA/Goddard/University of Arizona.
The authors note that a close approach to Earth that will occur in 2037 will be the next opportunity to collect radar data and therefore gauge the accuracy of their work while improving the trajectory projections even more. In their conclusion they add this (and note that we are dealing with not one but numerous gravitational keyholes)::
…improved orbital knowledge allowed us to refine the impact hazard assessment, which we extended through 2300. The dense structure of keyholes on the B-plane of the 2135 encounter with Earth (Chesley et al., 2014) made it unlikely to avoid all possible pathways to impact. Still, the uncertainties for the 2135 encounter decreased by a factor of about 20, and so many of the most significant impacts found by Chesley et al. (2014) are now ruled out.
The fact that we have a sample return on the way adds another plus for OSIRIS-REx. Dante Lauretta (University of Arizona) is principal investigator for the mission:
“The spacecraft is now returning home, carrying a precious sample from this fascinating ancient object that will help us better understand not only the history of the solar system but also the role of sunlight in altering Bennu’s orbit since we will measure the asteroid’s thermal properties at unprecedented scales in laboratories on Earth.”
The opportunity given us by OSIRIS-REx to test models and calculate future trajectory probabilities shows what can be done with objects with even a remote chance of striking the planet. The trajectory changes coming in 2135’s close approach will be watched carefully by way of further understanding how to tighten such calculations. By then, in the highly unlikely event of a dangerous deflection toward impact, we can hope to have methods in place to force a further trajectory change of our own.
The paper is Farnocchia et al., “Ephemeris and hazard assessment for near-Earth asteroid (101955) Bennu based on OSIRIS-REx data,” published online by Icarus 10 August 2011 (full text).
Habitability: Similar Magnetic Activity Links Stellar Types
Looking at flare activity in young M-dwarf stars, as we did in the last post, brings out a notable difference between these fast-spinning stars and stars like the Sun. Across stellar classifications from M- to F-, G- and K-class stars, there is commonality in the fusion of hydrogen into helium in the stellar cores. But the Sun has a zone at which energy carried toward the surface as radiative photons is absorbed or scattered by dense matter.
At this point, convection begins as colder matter moves downward and hot matter rises. This radiative zone giving way to convection is distinctive — stars in the M-class range, a third of the mass of the Sun and lower, do not possess a radiative core, but undergo convection throughout their interior.
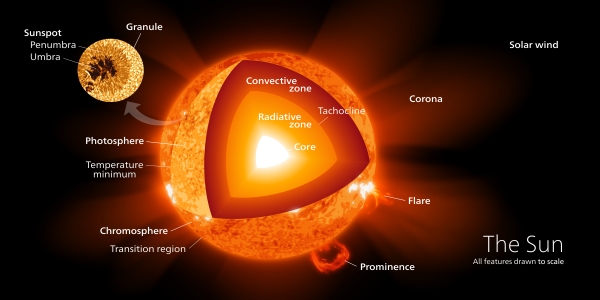
Image: Interior structure of the Sun. Credit: kelvinsong / Wikimedia Commons CC BY-SA 3.0.
If we’re going to account for magnetic phenomena like starspots, flares and coronal mass ejections, we can come up with a model that fits stars with a radiative core, but fully convective stars might be expected to have a different kind of magnetic dynamo. What stands out in the data, however, is that the relationship between the star’s rotation and its magnetic activity appears the same for stars on both sides of what I might call the ‘convective divide.’ In both cases, the magnetic dynamo seems to be efficient despite the fact that M-dwarfs are fully convective.
Digging further into the subject is a new paper out of Rice University, where modeling of these phenomena examines the linkage between the rotation of stars and the behavior of their surface magnetic flux. The flux in turn governs the luminosity of the star at X-ray wavelengths, giving us a way to probe magnetic activity and its potential effects on planets in these systems. The paper explaining the new model has just run in The Astrophysical Journal. Lead author Alison Farrish comments on the implications over time as rotation periods change:
“All stars spin down over their lifetimes as they shed angular momentum, and they get less active as a result. We think the sun in the past was more active and that might have affected the early atmospheric chemistry of Earth. So thinking about how the higher energy emissions from stars change over long timescales is pretty important to exoplanet studies.”
Image: Rice University scientists have shown that “cool” stars like the sun share dynamic surface behaviors that influence their energetic and magnetic environments. Stellar magnetic activity is key to whether a given star can host planets that support life. Credit: NASA.
Farrish and team used the Rossby number of our own star to model the behavior of other stars. This value measures stellar activity through the combination of rotational speed and subsurface liquid flows that influence how the magnetic flux is distributed on the stellar surface. Presumably the magnetic field in stars with a radiative zone is generated at the interface between the interior radiative region and the outer convective zone. The Rossby number relates the rotation of the star — determined through observation — to the internal convective activity of the star.
The results affirm that the mechanisms producing local ‘space weather’ are common across different stellar classes, meaning we can with some confidence examine planetary systems around M, F, G and K stars using the same model. The process generating a star’s magnetic field may thus turn out to be similar despite the presence, or lack, of a radiative core. Adds co-author Christopher Johns-Krull:
“A lot of ideas about how stars generate a magnetic field rely on there being a boundary between the radiative and the convection zones, so you would expect stars that don’t have that boundary to behave differently. This paper shows that in many ways, they behave just like the sun, once you adjust for their own peculiarities.”
Thus an M-dwarf with a Rossby number typical for its class shows magnetic behaviors close enough to the Sun for us to make predictions about their effect on its planets. The stellar magnetic field data are, in turn, affected by the activity cycle of individual stars, which the model does not include because this would demand lengthy observational study for each star. But from the perspective of magnetically active stars, the new model from Farrish and team can be applied to interactions within their systems.
We’re a long way from knowing whether M-dwarf systems like that at Proxima Centauri or the intriguing TRAPPIST-1 and L 98-59 could support living planets, but our models for their magnetic interactions can draw on what we see in our own Sun, despite its differences in age and stellar class. Refining that model for these systems will help us determine the most likely M-dwarf candidates for habitability.
The paper is Farrish et al., “Modeling Stellar Activity-rotation Relations in Unsaturated Cool Stars,” Astrophysical Journal Vol. 916, No. 2 (3 August 2021). Abstract.
Can M-Dwarf Planets Survive Stellar Flares?
We can learn a lot about stars by studying magnetic activity like starspots, flares and coronal mass ejections (CMEs). Starspots are particularly significant for scientists using radial velocity methods to detect planets, because they can sometimes mimic the signature of a planet in the data. But the astrobiology angle is also profound: Young M-dwarfs, known for flare activity, could be fatally compromised as hosts for life because strong flares can play havoc with planetary atmospheres.
Given the ubiquity of M-dwarfs — they’re the most common type of star in our galaxy — we’d like to know whether or not they are candidates for supporting life. A paper from Ekaterina Ilin and team at the Leibniz Institute for Astrophysics in Potsdam digs into the question by looking at the orientation of magnetic activity on young M-dwarfs.
The sample is small, though carefully chosen from the processing of over 3000 red dwarf signatures obtained by TESS, the Transiting Exoplanet Survey Satellite mission. The results are promising, indicating that the worst flare activity an M-dwarf can produce occurs along the poles of the star. If that is the case, then a young planetary system may remain unscathed. Here’s how Ilin describes this:
“We discovered that extremely large flares are launched from near the poles of red dwarf stars, rather than from their equator, as is typically the case on the Sun. Exoplanets that orbit in the same plane as the equator of the star, like the planets in our own solar system, could therefore be largely protected from such superflares, as these are directed upwards or downwards out of the exoplanet system. This could improve the prospects for the habitability of exoplanets around small host stars, which would otherwise be much more endangered by the energetic radiation and particles associated with flares compared to planets in the solar system.”

Image: Small stars flare actively and expel particles that can alter and evaporate the atmospheres of planets that orbit them. New findings suggest that large superflares prefer to occur at high latitudes, sparing planets that orbit around the stellar equator. Credit: AIP/ J. Fohlmeister.
This is provocative stuff, even if only four stars emerged from the TESS data as fitting the criteria the researchers were looking for. To understand how they winnowed these stars out, we need to look at PEPSI, the Potsdam Echelle Polarimetric and Spectroscopic Instrument, mounted at the Large Binocular Telescope (LBT) in Arizona. Feeding polarized light to the spectrograph, scientists using PEPSI have been able to use what is called the Zeeman effect — involving the polarization of spectral lines due to an external magnetic field — to analyze the field geometry of the field.
This earlier work has implied the existence of concentrated magnetic activity near the poles of fast rotating stars like young M-dwarfs, activity that emerges as spots and flares. While the Zeeman technique could reconstruct a stellar magnetic field, no observations of this polar clustering had previously been made — bear in mind that we cannot resolve the surface of the target stars. The Potsdam researchers were able to detect signs of polar clustering by analyzing white-light flares on their target stars, pinpointing the latitude of the flaring region from the shape of the light curve.
This works because modulations in brightness, caused by the young stars’ fast spin as the flare location rotates in and out of view on the stellar surface, carry useful information. M-dwarfs remain fast rotators much longer than stars like the Sun; in fact, the fast rotation enhances their magnetic and flare activity. The researchers were able to determine where on the star these flares occurred. According to the paper:
The exceptional morphology of the modulation allowed us to directly localize these flares between 55? and 81? latitude on the stellar surface. Our findings are evidence that strong magnetic fields tend to emerge close to the rotational poles of fast-rotating fully convective stars, and suggest a reduced impact of these flares on exoplanet habitability.
The kind of long-duration superflare activity considered most lethal for planetary atmospheres, in other words, occurs much closer to the pole than the weaker flares and spots found below 30?. On our own mature G-class star, sunspots and flares associated with them tend to occur near the equator. This paper offers, then, a continuing lifeline for those interested in the prospects for life around M-class stars, while also pointing to the need for what the authors call “the first fully empirical spatio-temporal flare reconstructions on low mass stars.” The emergence of such a model will help us draw broader conclusions on the impact of stellar magnetic activity on M-dwarf planets.
The paper is Ilin et al., “Giant white-light flares on fully convective stars occur at high latitudes,” accepted at Monthly Notices of the Royal Astronomical Society 05 August 2021 (abstract / Preprint). Thanks to Michael Fidler and Antonio Tavani for an early heads-up on this work.

Europa: Building the Clipper
Seeing spacecraft coming together is always exciting, and when it comes to Europa Clipper, what grabs my attention first is the radiation containment hardware. This is a hostile environment even for a craft that will attempt no landing, for flybys take sensitive electronics into the powerful radiation environment of Jupiter’s magnetosphere. 20,000 times stronger than Earth’s, Jupiter’s magnetic field creates a magnetosphere that affects the solar wind fully three million kilometers before it even reaches the planet, trapping charged particles from the Sun as well as Io.
We have to protect Europa Clipper from the intense radiation emerging out of all this, and in the image below you can see what the craft’s engineers have come up with. Now nearing completion at the Jet Propulsion Laboratory, the aluminum radiation vault will ultimately be attached to the top of the spacecraft’s propulsion module, connecting via kilometers of cabling that will allow its power box and computer to communicate with systems throughout the spacecraft. The duplicate vault shown below is used for stress testing before final assembly of the flight hardware.

Image: Engineers and technicians in a clean room at NASA’s Jet Propulsion Laboratory display the thick-walled aluminum vault they helped build for the Europa Clipper spacecraft. The vault will protect the spacecraft’s electronics from Jupiter’s intense radiation. In the background is a duplicate vault. Credit: NASA/JPL-Caltech.
The ATLO phase (Assembly, Test, and Launch Operations) begins in the spring of 2022 at JPL, with the radiation vault being one of the first components in place as Europa Clipper enters its final stage of fabrication. The 3-meter tall propulsion module was recently moved from Goddard Space Flight Center in Greenbelt, Maryland to the Johns Hopkins Applied Physics Laboratory (APL) in preparation for the installation of electronics, radios, antennae and cabling. Science instruments, meanwhile, are being tested at the universities and other institutions contributing to the mission.
Jan Chodas (JPL) is Europa Clipper Project Manager:
“It’s really exciting to see the progression of flight hardware moving forward this year as the various elements are put together bit by bit and tested. The project team is energized and more focused than ever on delivering a spacecraft with an exquisite instrument suite that promises to revolutionize our knowledge of Europa.”
Before the ATLO phase begins, Europa Clipper will also be subject to a System Integration Review later this year, a process in which all instruments are inspected and plans for the assembly and testing of the spacecraft are finalized. The destination for all these components and instruments is the main clean room at JPL in Pasadena, where what NASA describes as the ‘choreography’ of building a flagship mission will draw together components from workshops and laboratories in the US and Europe.

Image: Contamination control engineers in a clean room at NASA’s Goddard Space Flight Center in Greenbelt, Maryland, evaluate a propellant tank before it is installed in NASA’s Europa Clipper spacecraft. The tank is one of two that will be used to hold the spacecraft’s propellant. It will be inserted into the cylinder seen at left in the background, one of two cylinders that make up the propulsion module. Credit: NASA/GSFC Denny Henry.
The travels of the propulsion module emphasize the collaborative nature of any complex spacecraft assembly. The two cylinders making up the module were built at the Applied Physics Laboratory and then shipped to JPL, where thermal tubing carrying coolant to regulate the spacecraft’s temperature in deep space was added. The cylinders then went to Goddard, where the propellant tanks were installed inside them and the craft’s sixteen rocket engines were attached to the outside. It then returned to APL for the installation of electronics and cabling mentioned above.

Image: Engineers and technicians in a clean room at NASA’s Goddard Space Flight Center in Greenbelt, Maryland, integrate the tanks that will contain helium pressurant onto the propulsion module of NASA’s Europa Clipper spacecraft. Credit: NASA/GSFC Barbara Lambert.
Connecting to the thermal tubing will be Europa Clipper’s radiator, which will radiate enough heat into space to keep the spacecraft in its operating temperature range. APL is now integrating the propulsion module and the radios, antennae and cabling for communications, while a company called Applied Aerospace Structures Corporation in Stockton, California is building the 3-meter high-gain antenna. By the spring of next year, the antenna will be in place at JPL for insertion in the ATLO process.
Nine science instruments will fly aboard Europa Clipper, all being assembled and undergoing testing at NASA centers as well as partner institutions and private vendors. The spacecraft is to investigate the depth of the internal ocean as well as its salinity and the thickness of the ice crust. The latter is obviously a huge factor in any future plans to sample the ocean beneath the ice, but so is the question of whether Europa vents subsurface water into space through plumes that may one day be sampled.

Image: NASA’s Jet Propulsion Laboratory in Southern California is building the spectrometer for the agency’s Europa Clipper mission. Called the Mapping Imaging Spectrometer for Europa (MISE), it is seen in the midst of assembly in a clean room at JPL. Pronounced “mize,” the instrument will analyze infrared light reflected from Jupiter’s moon Europa and will map the distribution of organics and salts on the surface to help scientists understand if the moon’s global ocean – which lies beneath a thick layer of ice – is habitable. Credit: NASA/JPL-Caltech.
We’ll finally be able to update those Galileo images that have served scientists so well in the study of Europa’s surface with new, detailed looks at the surface geology. Launch is currently planned for October, 2024 aboard a Falcon Heavy rocket. Europa Clipper isn’t a life detection mission, but we’ll learn a good deal more about Europa’s potential for supporting life. What kind of mission grows out of that is something it would be foolhardy to predict. One step at a time as Europa reveals its mysteries.
L 98-59 b: A Rocky World with Half the Mass of Venus
ESPRESSO comes through. The spectrograph, mounted on the European Southern Observatory’s Very Large Telescope, has produced data allowing astronomers to calculate the mass of the lightest exoplanet ever measured using radial velocity techniques. The star is L 98-59, an M-dwarf about a third of the mass of the Sun some 35 light years away in the southern constellation Volans. It was already known to host three planets in tight orbits of 2.25 days, 3.7 days and 7.5 days. The innermost world, L 98-59b, has now been determined to have roughly half the mass of Venus.
What extraordinary precision from ESPRESSO (Echelle SPectrograph for Rocky Exoplanets and Stable Spectroscopic Observations). The three previously known L 98-59 planets were discovered in data from TESS, the Transiting Exoplanet Survey Satellite, which spots dips in the lightcurve from a star when a planet crosses its face.
Adding ESPRESSO’s data, and incorporating previous data from HARPS, has allowed Olivier Demangeon (Instituto de Astrofísica e Ciências do Espaço, University of Porto) and team to refine the planets’ mass. Because we already know their radii through transits, we can constrain the density of these rocky worlds. Intriguingly, 30% of L 98-59 d’s mass could be water.
What stands out here, though, is the confirmation of ESPRESSO’s capabilities as we continue to drill down into the centimeters-per-second range that will allow us to probe small rocky worlds around other stars. We’ve seen rapid growth in spectrography through ESPRESSO as well as NEID and, of course, HARPS (High Accuracy Radial Velocity Planet Searcher), which has long been in the forefront of the exoplanet hunt at ESO’s 3.6m telescope at La Silla Observatory in Chile.
ESPRESSO continues to push the boundaries of radial velocity planet detection. There is no hyperbole at all in the conclusion to the paper on this work, which notes that the refinement of mass for the planets in this system, particularly the innermost world:
…represents a new milestone which illustrates the capability of ESPRESSO to yield the mass of planets with RV signatures of the order of 10 cm s-1 in multi-planetary systems even with the presence of stellar activity.
The ESPRESSO data also flag a fourth planet around this star, along with hints of a possible fifth, the latter of which would be in the star’s liquid water habitable zone. The detected planet e has an orbital period of 12.80 days with a minimum mass of 3 Earth masses, while the candidate fifth planet has a period of 23.2 days and a minimum mass of 2.46 Earth masses. It would be in the star’s habitable zone and thus of high interest if confirmed, although there remains the possibility that the signal in the data is the result of stellar activity.
There are no signs of transits from either of these worlds. As this system is likely to become a benchmark for planetary analysis in nearby systems, we’ll keep an eye on the confirmation process for the planet candidate here.

Image: Comparison of the L 98-59 exoplanet system with the inner Solar System.
The three inner worlds at L 98-59 are candidates for atmospheric study through transmission spectrography, where astronomers examine light from the star as filtered through a planetary atmosphere during a transit. The astronomers note that in addition to potential analysis via the James Webb Space Telescope, the Extremely Large Telescope under construction in Chile’s Atacama Desert — scheduled to begin observations in 2027 — may be able to study the atmospheres of these planets from the ground.
In any event, further work with ESPRESSO, the Hubble Space Telescope, and future observatories like NIRPS (Near Infra Red Planet Searcher in Chile) and the Ariel space telescope (Atmospheric Remote-sensing Infrared Exoplanet Large-survey) should be available for atmospheric studies in this interesting system. Adds Demangeon:
“This system announces what is to come. We, as a society, have been chasing terrestrial planets since the birth of astronomy and now we are finally getting closer and closer to the detection of a terrestrial planet in the habitable zone of its star, of which we could study the atmosphere.”
An additional note relates to the tightness of planetary system configurations in multiple planet systems. This is from the paper’s conclusion:
According to exoplanet archive (Akeson et al. 2013), we currently know 739 multi-planetary systems. A large fraction of them (~ 60%) were discovered by the Kepler survey (Borucki et al. 2010; Lissauer et al. 2011). From a detailed characterization and analysis of the properties of the Kepler multiplanetary systems, Weiss et al. (2018, hereafter W18) extracted the “peas in a pod” configuration. They observed that consecutive planets in the same system tend to have similar sizes. They also appear to be preferentially regularly spaced. The authors also noted that the smaller the planets, the tighter their orbital configuration is… [W]e conclude that the L 98-59 system is closely following the “peas in a pod” configuration…
A useful fact, and one that, as the authors add, “further strengthens the universality of this configuration and the constraints that it brings on planet formation theories.”
The paper is Demangeon et al., “Warm terrestrial planet with half the mass of Venus transiting a nearby star,” accepted at Astronomy & Astrophysics (abstract).

Follow with RSS or E-Mail
